
Бесплатный фрагмент - Боль в спине. За кулисами лечения и реабилитации
Часть III. Механизмы «неспецифической» боли


ГЛАВА 5. БЕГЛО ПО ЭВОЛЮЦИИ ЧЕЛОВЕКА
Вероятно, многим знакомо такое выражение, как «остеохондроз является расплатой за прямохождение». Человек как отдельный вид млекопитающего прошел достаточно долгий эволюционный путь, в результате которого приобрел привычный всем вид. И, если, антропологам, специалистам по биомеханике и ортопедам все более-менее ясно относительно «расплаты за прямохождение», то для других специалистов, принимающих участие в лечении дегенеративных заболеваний позвоночника, многие вопросы остаются до конца не понятыми. Мало кто может внятно объяснить что-либо вообще по данному вопросу. И, как следствие, в ряду этиологических (являющихся причиной) факторов развития дегенеративно-дистрофических процессов эволюционный фактор остается наиболее игнорируемым среди рядовых врачей. Роль/влияние данного фактора недооценивают, наивно полагая, что за 4 млн. лет эволюции человек приобрел оптимальную структуру/модель двигательного аппарата. Большинство обывателей вообще не знакомы с данной проблемой или считают ее сугубо научной. Это заблуждение, за которое часто приходиться дорого платить. И речь идет не только о заболеваниях позвоночника, но и о других смежных заболеваниях опорно-двигательного аппарата.
С переходом к прямохождению человек приобрел множество предпосылок к заболеваниям, напрямую связанных с передвижением в ортоградном (вертикальном) положении. В данном случае речь идет именно о предпосылках, а не о факторах, вызывающих данные заболевания.
Наиболее распространенные:
— сложная родовая деятельность*, требующая родовспоможения (зависит от типа таза);
— остеопороз позвоночника, вызванный снижением минерализации в вертикальном поло-жении, приводящий к компрессионным переломам, особенно у женщин;
— сколиотические осанки, идиопатические и дегенеративные сколиозы;
— варикозное поражение вен нижних конечностей;
— дегенеративно-дистрофическое поражение позвоночника;
— старческая миелопатия на шейном уровне;
— дегенеративные поражения суставов (коксартрозы, гонартрозы).
Выражение «Расплата за прямохождение» звучит как приговор. Автор фразы был не в настроении и явно сгустил краски. Скорее абонентская плата за использование более совершенной эволюционной модели. На самом деле прямохождение — это дар человечеству, за который мы должны благодарить судьбу, волю случая или эволюцию.
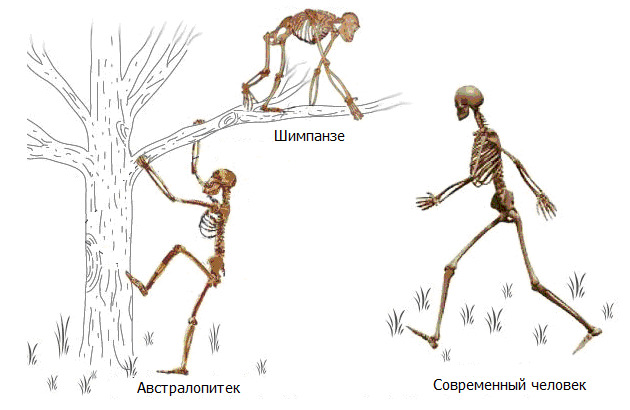
Привычное всем прямохождение. За кажущейся простотой фразы стоит колоссальный эволюционный скачок, который привел к фундаментальному изменению в способе передвижения предка человека. Переход к прямохождению (ортоградное положение — т.е. вертикально) не просто снизил энергетические затраты, уменьшив участие мышечной системы в акте передвижения, но и освободил передние конечности от необходимости нести опорную статодинамическую нагрузку. Со временем освобожденные передние конечности эволюционировали в ловкие манипуляторы, давшие мощный толчок к развитию мозга.
Человек уникален тем, что он является единственным млекопитающим, которое передвигается вертикально и, преимущественно, использует только две нижние конечности. Читатель может возразить, что многие животные при желании свободно переходят в вертикальное положение и довольно свободно передвигаются. Да, это так. Но, вопрос в энергетической стоимости такой «ходьбы» и приспособленности скелета для длительного поддержания вертикального положения. Человек также может передвигаться на четвереньках, но насколько это удобно — судить каждому в отдельности.
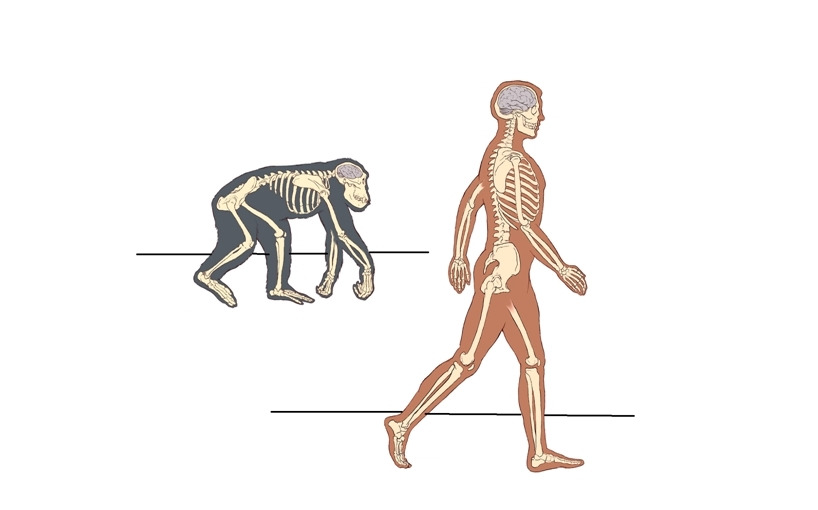
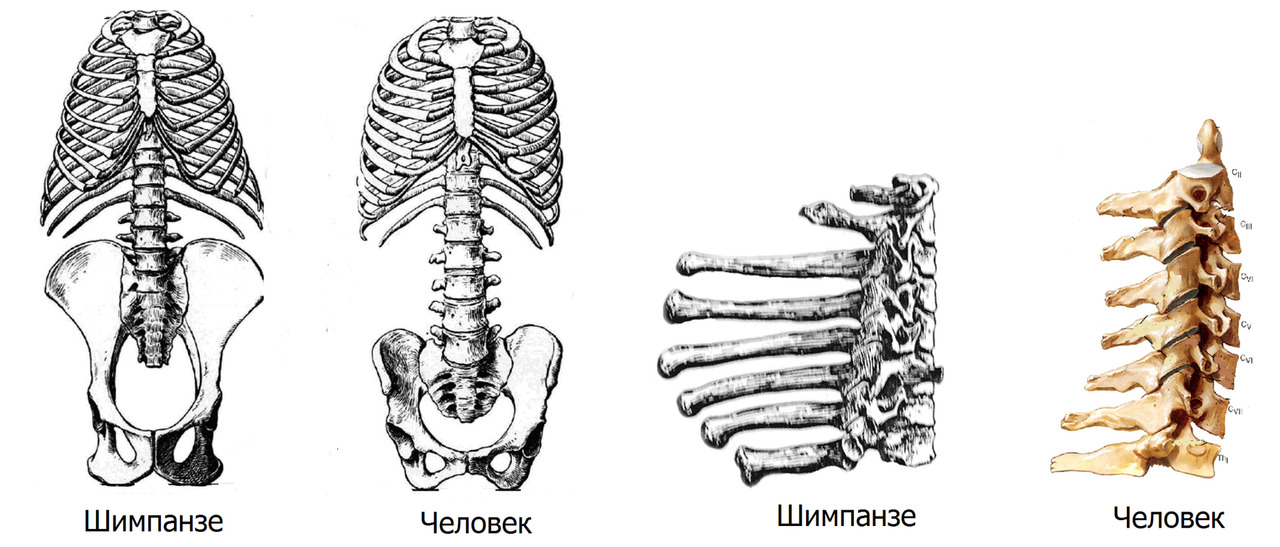
Чтобы не влезать в дебри современной антропологической теории о происхождении человека, сравнение проведем не с древним общим предком человека и современных обезьян, а с ближайшим родственником человека — карликовым шимпанзе Бонобо.
Предок предка (это не тавтология; в контексте описываемой проблемы это важное разграничение) современного человека большую часть своей жизни проводил на деревьях с эпизодами передвижения по земле. Перемещение осуществлялось на четырех конечностях с кратковременными периодами «ходьбы» на задних лапах. С точки зрения человеческой биомеханики, передние и задние конечности у шимпанзе — это четыре руки. Задние конечности хоть и имеют отличие от передних, но, все-таки, это задние руки. Строение скелета и позвоночника общего предка приспособлено для передвижения вдоль площади опоры способом чередования толчки-отталкивания. По деревьям — комбинация подтягиваний-отталкиваний. То есть такой тип передвижения, который требует больших мышечных усилий с двух пар конечностей.

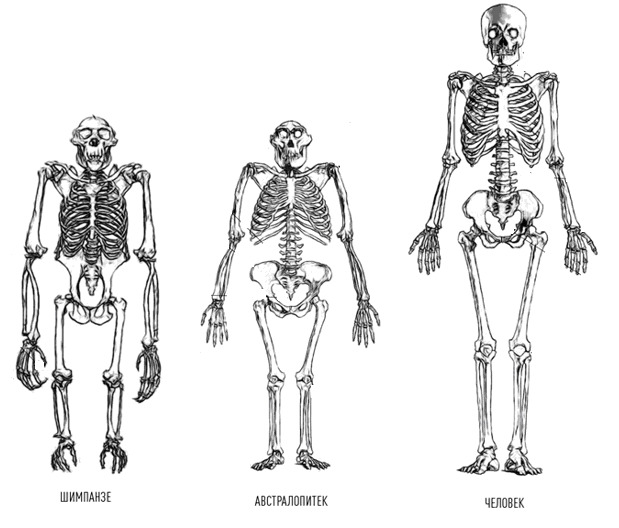
Беглый сравнительный анализ поможет выделить особенности в строении скелета шимпанзе и человека, которые являются предпосылкой для будущих проблем с позвоночником у человека.
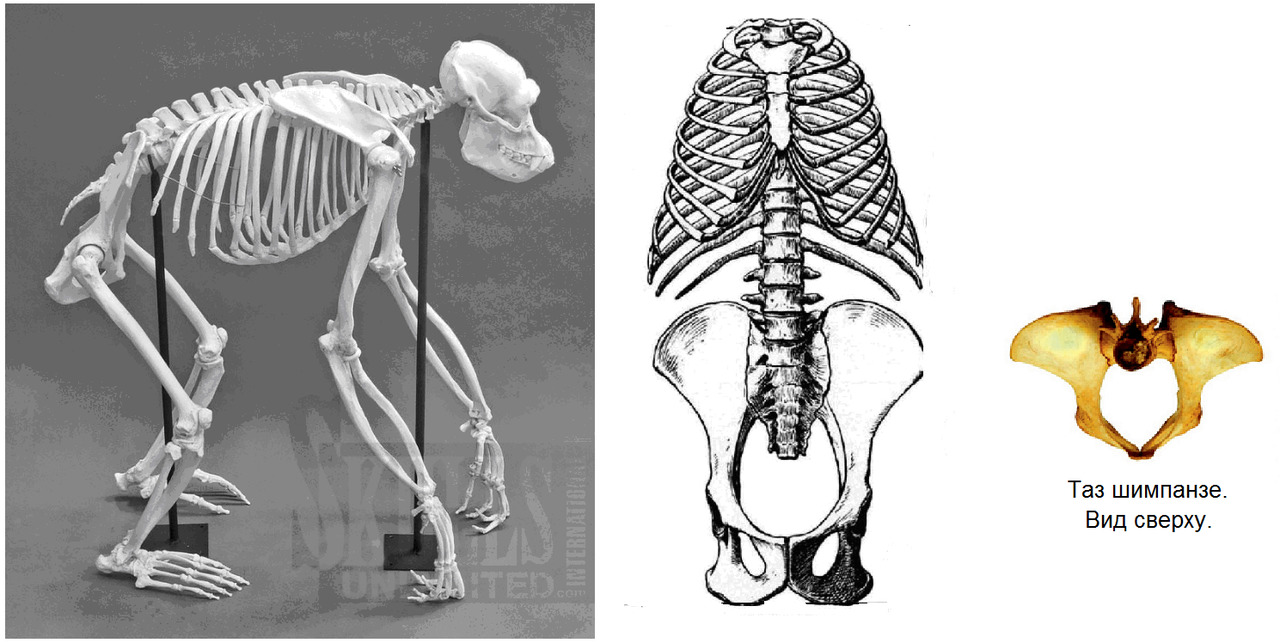
Шимпанзе: позвоночник выгнут легкой дугой, таз длинный, узкий, плоский. Крестец узкий, длинный, высоко расположен в тазу относительно головок бедренных костей. Вертлужная впадина мелкая. Крыша вертлужной впадины не выражена. Крылья подвздошных костей расположены преимущественно во фронтальной плоскости. Грудная клетка конусообразная.
Нижние ребра близко расположены к крыльям подвздошных костей. Позвоночник жесткий. Движения головы ограничены жесткостью выйной связки и длиной остистых отростков шейных позвонков.
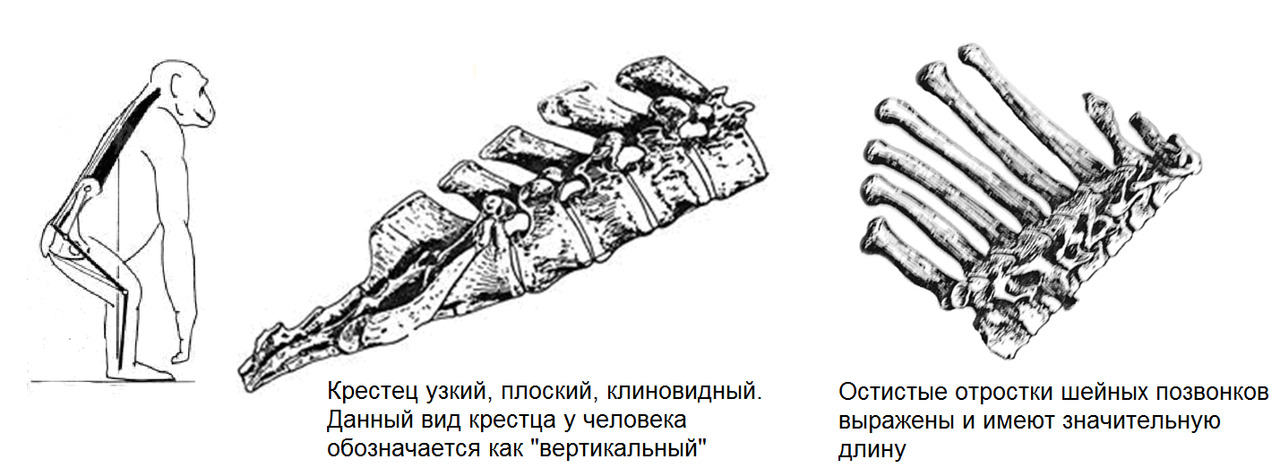
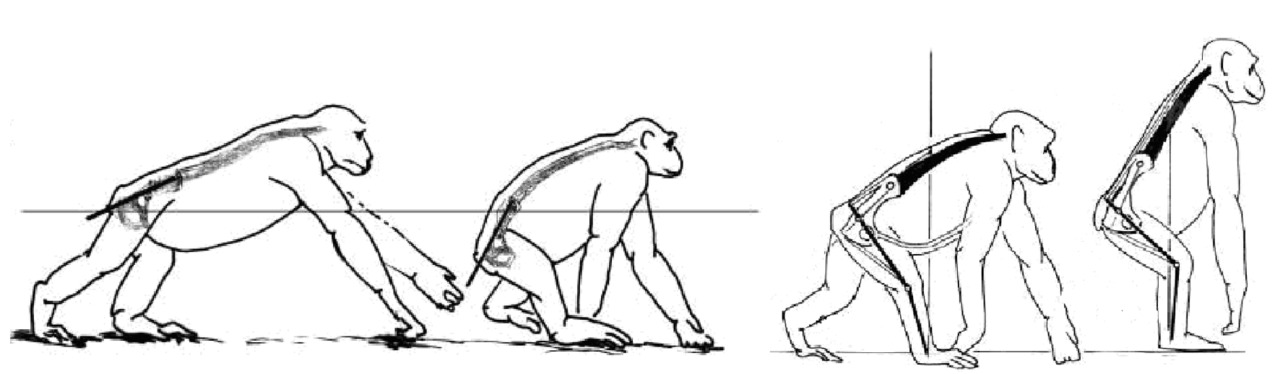
Чтобы принять относительно «вертикальное» положение, шимпанзе должен стоять на согнутых задних лапах с некоторым наклоном корпуса вперед. Такое положение не может поддерживаться длительно по причине перенапряжения мышц спины и нижних конечностей. Также оно энергозатратно и приводит к быстрому утомлению. Ходьба возможна только «вразвалку» по типу «перекантовывания» всего тела. Общий центр тяжести находится высоко относительно головок бедренных костей и смещен вперед относительно позвоночника. Такая ходьба требует дополнительных усилий на балансировку, что еще больше повышает энергозатраты. Поэтому такой вид передвижения используется в крайних случаях.
В процессе эволюционного перехода от перемещения на четырех конечностях к ходьбе на двух ногах, наиболее радикальные изменения произошли в поясе задних конечностей.
Для перехода пояса задних конечностей примата в пояс нижних конечностей предка человека потребовались коренные изменения в таких звеньях скелета:
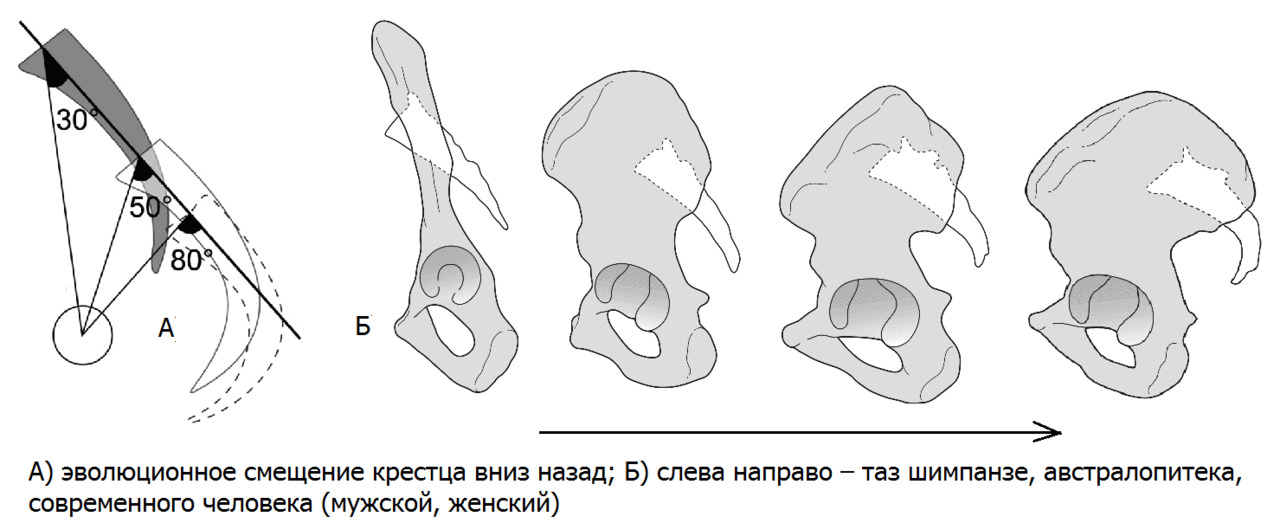
— крестец стал шире, короче, приобрел изогнутую форму и стал более массивным.
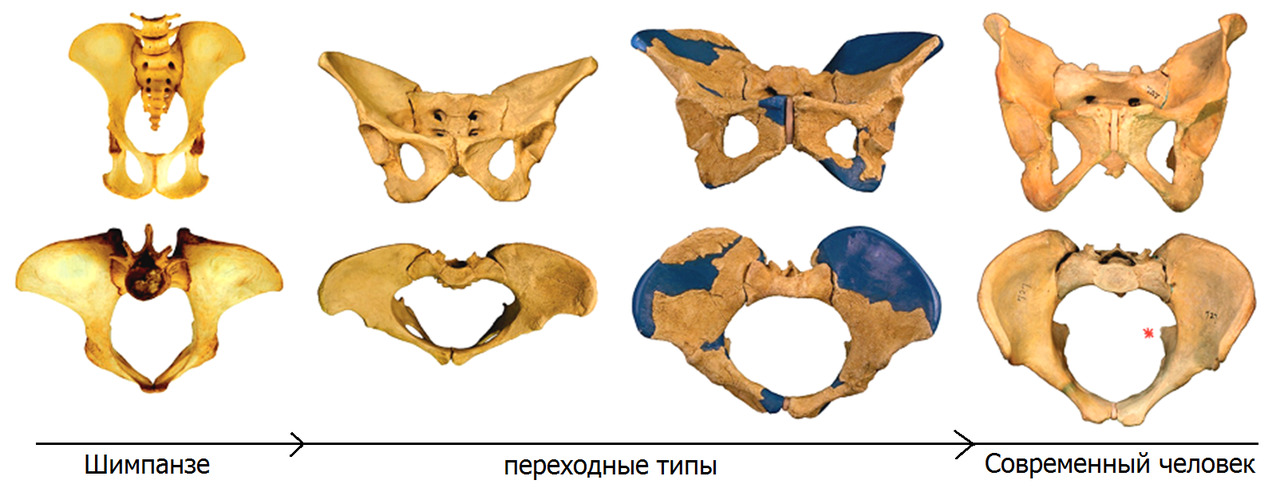
Таз из формы «лопаты» приобрел форму «чаши». Для улучшения балансировки туловища относительно тазобедренных суставов, крестец опустился вниз и занял более горизонтальное положение;
— подвздошные кости стали короче, шире, массивней и совершили поворот из фронтальной плоскости в сагиттальную. Раскрытие подвздошных костей в сторону привело к увеличение опорной пощади верхней апертуры таза в аксиальной плоскости.
— значительно увеличился диаметр и глубина вертлужной впадины. Суставная впадина тазобедренного сустава приобрела верхний свод — крышу, тем самым увеличив площадь соприкосновения с головкой бедренной кости
— бедренная кость стала длинней, массивней, увеличился размер головки бедра. Увеличение длины шейки бедра увеличило длину плеча рычага малой и средней ягодичной мышцы, которые переместились вместе с разворотом подвздошной кости в сагиттальную плоскость. Такой механизм позволил компенсировать боковой наклон во время ходьбы при одноопорной фазе шага.
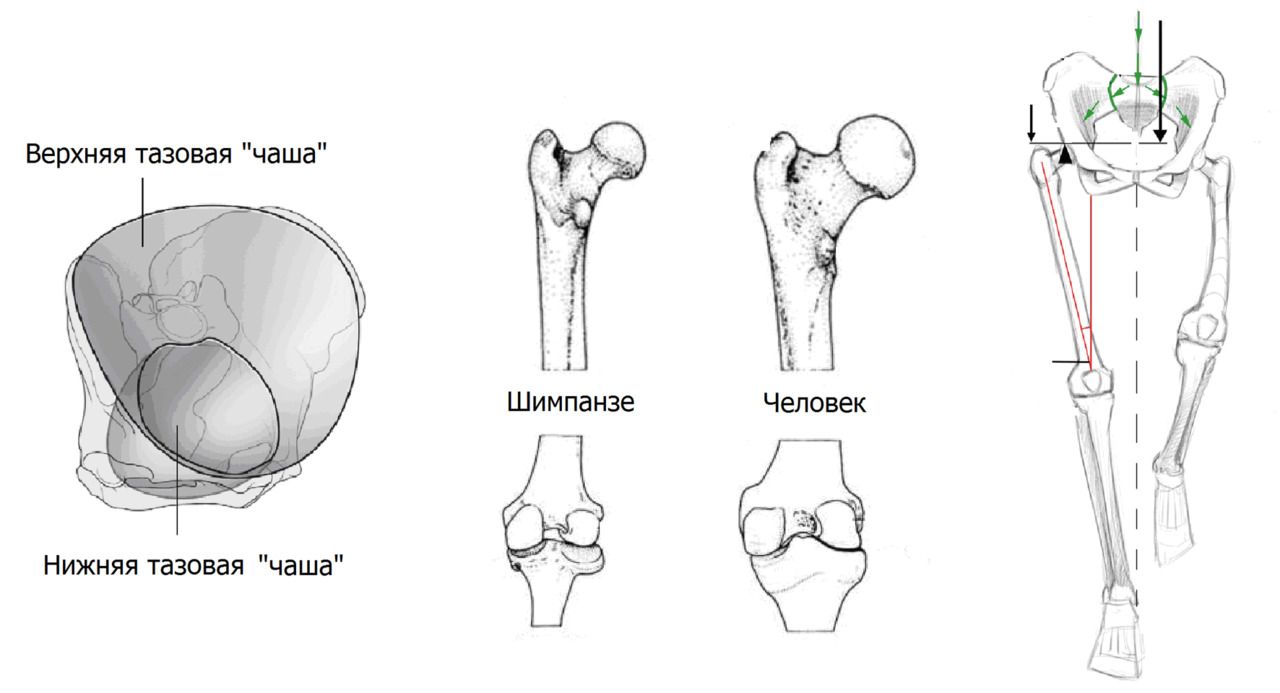
— значительно увеличился размер медиального мыщелка и глубина подчашечной вырезки, что усилило стабилизацию коленного сустава.
— голеностопный сустав утратил возможность поворота стопы. Пяточная кость стала массивной и увеличилась по длине, обеспечив рычаг мышцам разгибателям голени.
«Хватающая» стопа приобрела полноценную функцию опорной стопы: большой палец приведен к остальным и фиксирован общим единым комплексом связок. Кости плюсны/предплюсны и фаланг увеличились в размере. Стопа приобрела три свода, составляющие опорный треугольник стопы. Стопа утратила «хват», но приобрела функцию амортизатора. Сложная система глубоких связок, фасций, мышц стопы и голени определила стопу как рецепторное поле, являющееся основным источником сигналов с опоры.
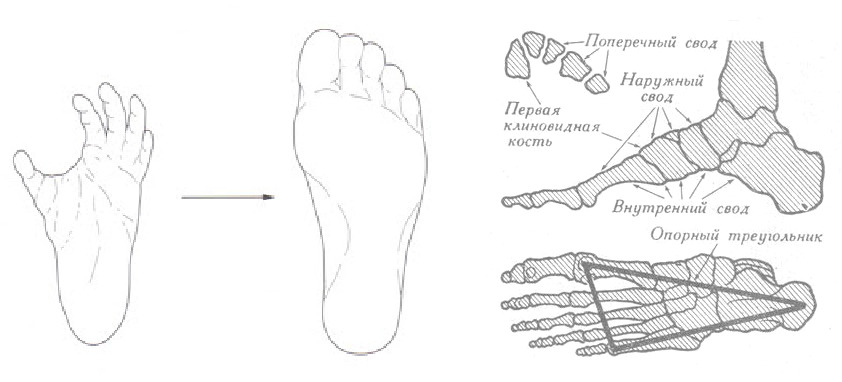
В целом вся нижняя конечность относительно таза заняла вальгусное положение — под углом к вертикальной оси. Выраженный вальгус (Х форма) нижней конечности позволил при ходьбе выносить ногу максимально близко к проекции общего центра масс (общего центра тяжести) компенсируя расшатывание по сторонам.
Выигрыш в биомеханике привел к значительной потере прочности конструкции.
Условные значимые потери
Необходимость удерживать общий центр тяжести с минимальными энергозатратами непосредственно над опорной частью — головками бедренных костей, привел к «централизации» поясничного отдела позвоночника путем формирования прогиба вперед в поясничной области — лордозированию. Это достигнуто благодаря формированию клиновидных тел позвонков и дисков с частичной потерей длины остистых отростков в пояснично-крестцовом переходе. Потеря длины отростка — это потеря длины рычага, через который передается мышечная тяга.
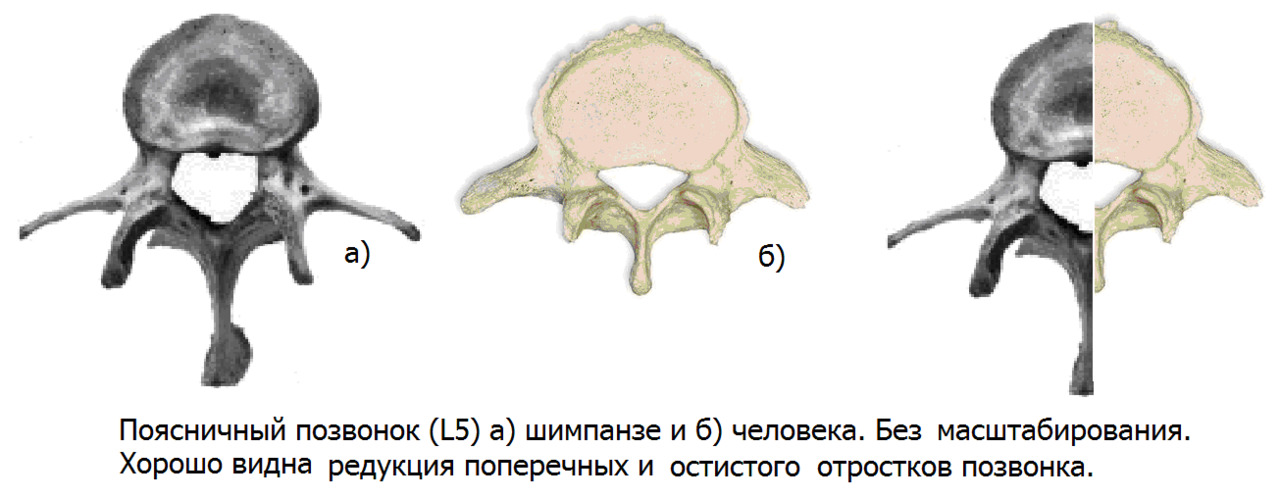
Также поперечные отростки поясничных позвонков утратили треть своей длины. Такое резкое уменьшение длины рычага с одномоментной выраженной потерей мышечной массы в поясничном отделе позвоночника привело к предрасположенности к ротационной нестабильности в поясничном отделе позвоночника. Прежде всего, это связано с косой установкой опорной площадки 1 крестцового позвонка относительно горизонтальной плоскости. Чем больше угол между замыкательной пластинкой 1 крестцового позвонка и горизонтальной плоскостью, тем выше крутящий момент на поясничный отдел позвоночника при разнице длины ног или мышечном дисбалансе.
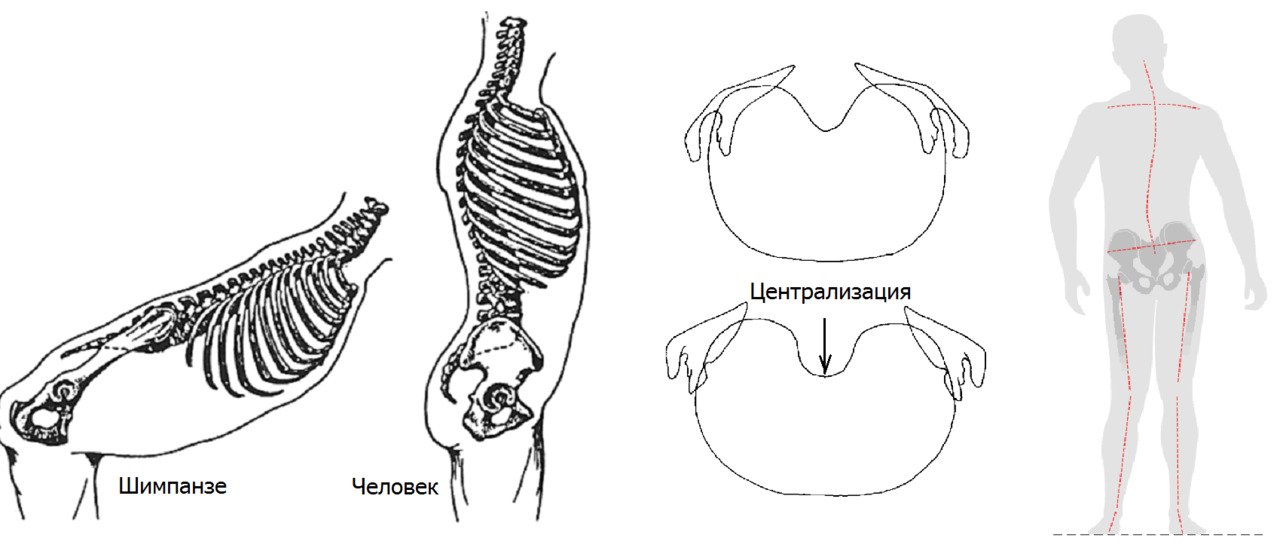
Комбинация централизации (лордозирование) с незначительным перекосом таза и приобретенная предрасположенность к ротации в поясничном отделе, приводят к формированию сколиотической дуги поясничного отдела позвоночника с вращением позвонков.
Искривление в поясничной области запускает волну/цепь компенсаторных искривлений, сопровождаемых вращением позвонков в противоположную сторону на грудном и шейном уровнях — сколиозирование.
Формирующийся грудной горб приводит к нарушению функции ребер и снижению внешнего дыхания. Шейный сколиоз нарушает работу суставов головы, значительно искажая проприоцепцию, чем ухудшает работу вестибулярной системы.
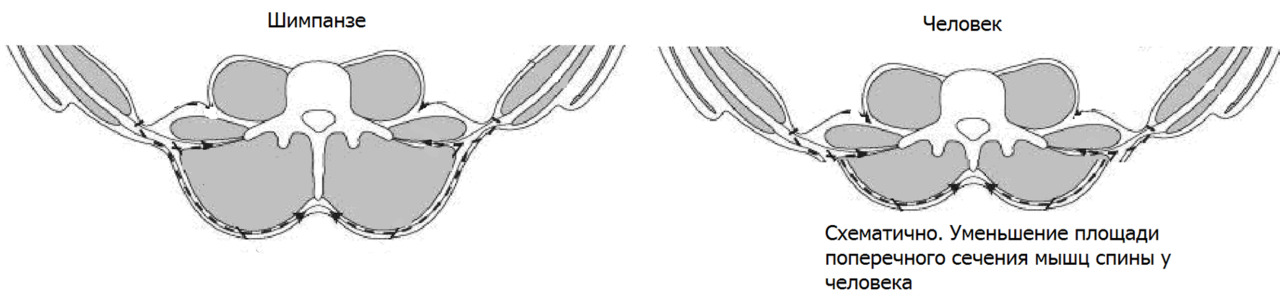
Снижение объема мышечной массы с сопутствующим снижением связочного компонента на поясничном уровне и на уровне пояснично–крестцового перехода привело к предрасположенности к развитию ранней гипермобильности/нестабильности данного региона.
Изменения также коснулись формы и направления суставных отростков. На поясничном уровне они совершили разворот из фронтальной плоскости в сагиттальную, что обеспечило возможность большего прогиба в поясничном отделе. У современного человека на уровне пояснично-крестцового перехода могут встречаться оба варианта расположения суставов и зависит от типа таза.
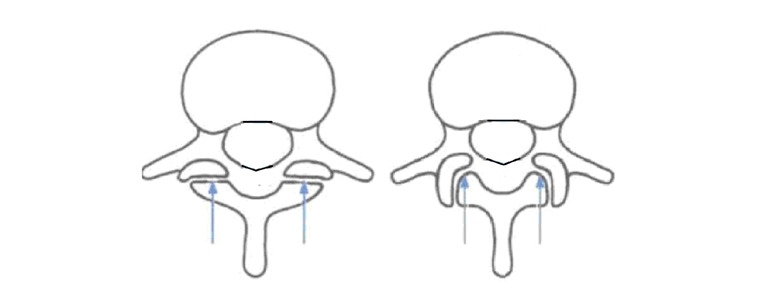
Значительное изменение произошло в форме позвоночного канала на нижнепоясничном уровне. Из формы круга или овала он трансформировался в треугольник. Компенсаторное увеличение опорной площади позвонков сверху вниз привел к сужению позвоночного канала на протяжении всего поясничного отдела позвоночника, резко сузив резервные пространства, тем самым ограничив, а у некоторых индивидуумов сведя к минимуму компенсаторные возможности организма при дегенеративных процессах в ПДС.
Необходимость компенсирующей противоротации грудной клетки относительно таза во время ходьбы привела к уменьшению нижних ребер и изменению формы и функции позвонков, лежащих на границе перехода грудной клетки и поясничного отдела. Форма грудной клетки из конусовидной стала бочкообразной.
Необходимость слежения за угрозой сверху и фиксации взгляда в горизонтальной плоскости привела к формированию шейного прогиба вперед — лордоза. Такое новшество произошло за счет значительного укорочения длины остистых отростков, их наклону вниз и расщеплению.
Также произошла значительная потеря мышечной массы шеи аналогично поясничному отделу. Это значительно увеличило подвижность и возможность смотреть вверх разгибая шею, при этом стоя на выпрямленных ногах.
В сумме имеем выраженное тройное переразгибание в звеньях скелета: бедро переразогнуто относительно таза, поясничный отдел — относительно таза, шейный отдел — относительно грудной клетки.
Большое затылочное отверстие сместилось в центр основания черепа, увеличив длину плеча рычага затылочной кости, что увеличило эффективность подзатылочных мышц и мышц надплечья. Череп относительно шейных позвонков отбалансирован формой за счет выраженного увеличения затылочной части черепа. Таким образом, голова в сагиттальной плоскости стала более сбалансированной по отношению к шее и тазу, но в плане фиксации механически «проиграв» в прочности и надежности конструкции из-за потери объема мышечной и связочной ткани.
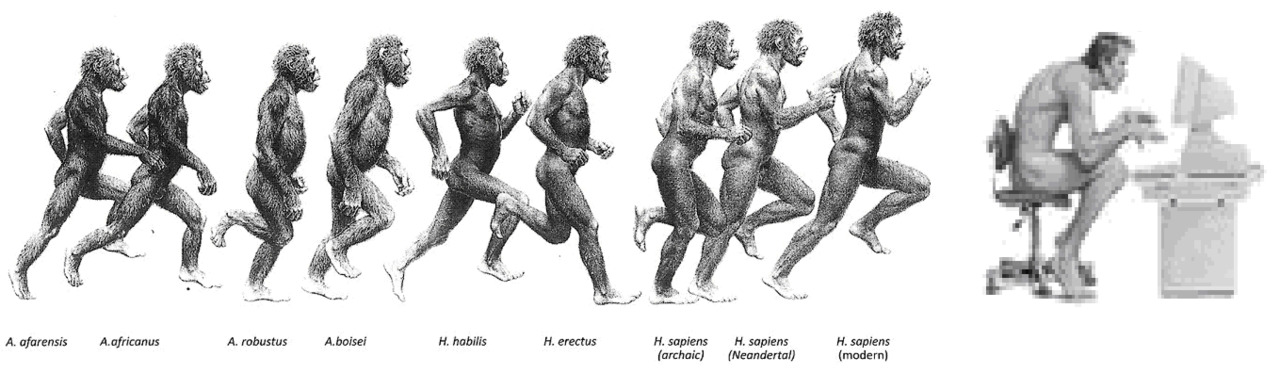
Эволюция человека шла в два основных этапа. На первом промежуточном этапе древние гоминиды могли передвигаться как привычным для нас способом — вертикально, так и успешно лазить по деревьям в случае необходимости. Ископаемые останки промежуточных типов гоминидов указывают, что Австралопитек равноценно использовал оба типа передвижения: на двух ногах, если приходилось пересекать открытые местности, и использовать четыре конечности во время укрытия на деревьях от нападения хищников или при сборе пропитания. Резкая смена климата и переход к кочевому способу жизнедеятельности на открытой местности окончательно привела к полной адаптации к прямохождению как основному способу передвижения. Необходимость мигрировать в места обитания с малым количеством лесов и протяженными равнинами привела к выработке дополнительных эволюционных изменений — выраженному изменению формы стопы, что облегчило ходьбу.
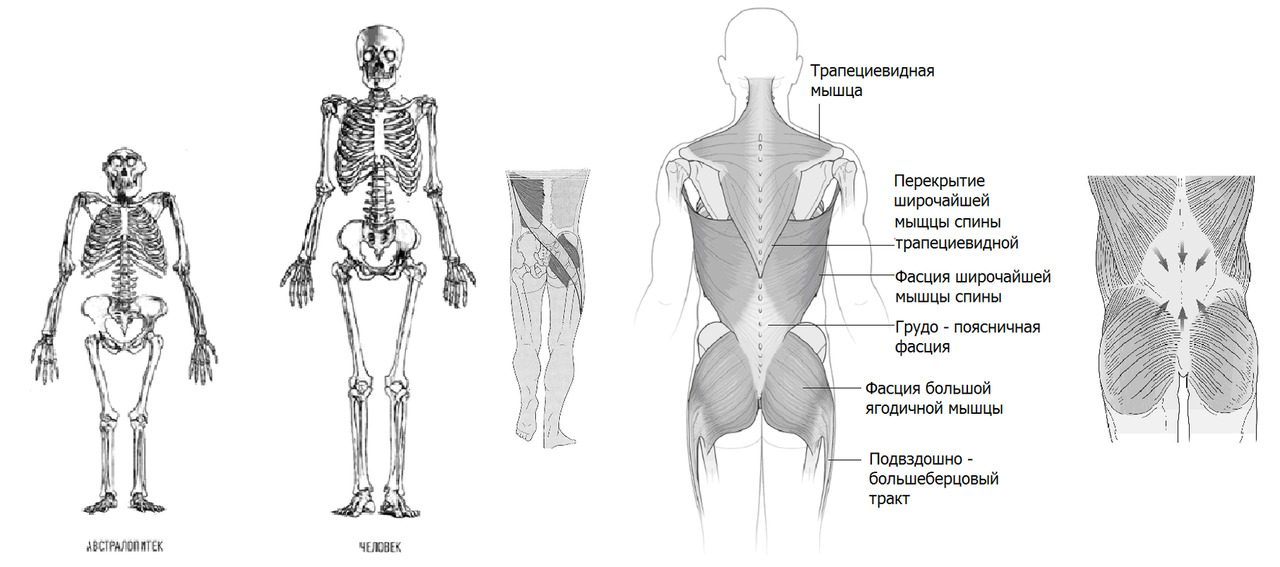
При переходе с частичного на полное вертикальное передвижение произошла значительная выраженная редукция двух мышечных групп, ответственных за лазанье по деревьям и ходьбы на четвереньках: широчайшей мышцы спины и больших ягодичных мышц. Широчайшая мышца спины в локомоции не используется и у современного человека выражена только у лиц, занимающихся определенными видами спорта. С большой ягодичной та же ситуация: данная мышца работает только как мощный разгибатель бедра (фаза толчка), одновременно препятствуя падению корпуса вперед при быстром беге и как стабилизатор/корректор положения противоположного бедра при сгибании (фаза выноса ноги). Это мышца быстрого бега: догнать жертву или уйти от преследования. Или при толчке при крутом подъеме вверх. Аналог взбирания по дереву. Другими словами, данная мышца может быть достаточно развита только у спортсменов с определенными двигательными специализациями [легкоатлеты — спринтеры]. К слову, наличие больших жировых подушек на ягодицах не заменяет сильные выносливые мышцы.
Редукция или выраженное снижение силы двух упомянутых мышечных групп в определенной мере является критичной для функции поясничного отдела позвоночника. Современный человек функционально утратил силу как раз именно тех мышц, которые вносят существенный вклад в стабилизацию поясничного отдела. Чисто анатомиически данные мышцы еще сохранились, но функционально они уже мало к чему пригодны без специальной тренировки (попробуйте подтянуться 50 раз на перекладине). Крестообразно пересекаясь, волокна этих мышц формируют надосную связку позвоночника и натягивают пояснично-грудную фасцию, которая является основным стабилизатором поясничного отдела позвоночника. Значительная потеря мышечной массы позвоночника при переходе от Бонобо к Австралопитеку, затем потеря двух стабилизирующих мышечных групп при переходе от Австралопитека к Человеку прямоходящему значительно ослабили всю конструкцию позвоночного столба и поясничного отдела в частности.
Переход от Австралопитека к Человеку прямоходящему ознаменовался еще одним значительным изменением — увеличением длины нижней конечности за счет увеличения длины бедренной и берцовых костей.
Вместе с тем, для снижения инерции верхнего сегмента произошло значительное укорочение верхних конечностей, которые теперь стали выполнять функцию балансира и ротационного стабилизатора корпуса, компенсируя тем самым крутящий момент при ходьбе и беге. Для снижения инерционной составляющей пришлось пожертвовать не только мышечной массой передних конечностей, но и мышечной массой всего туловища, включая мышцы спины. Также значительно снизилась минерализация костей, тем самым максимально облегчив скелет туловища. Общий центр масс сместился максимально вниз к головкам бедренных костей. Такое смещение ОЦМ уравновесило инерционные моменты сил верхней части — корпуса и нижних конечностей, обеспечив возможность балансировки тела с минимальными мышечными затратами и при минимальной площади опоры. Смещение ОЦМ в таз «замкнуло» позвоночник на бедре, убрав необходимость мышечного удержания равновесия большими мышечными группами.
Снятие нагрузки с рук привело к двойной нагрузке на нижние конечности. Сустав бедра начал работать не только как консольная балка, но и как несущая опора. Межпозвонковые диски, до этого работавшие только в режиме буфера с чередованием фаз растяжение — компрессия с элементами ротации, теперь стали выполнять не свойственную им функцию: функция буфера (работа в горизонтальной плоскости) сменилась на функцию амортизации (работа в вертикальной плоскости). Но самым важным изменением в данной конструкции явилось то, что в вертикальном положении диск начал выполнять функцию точки опоры рычага первого рода вышележащего сегмента.
Если грубо сравнивать биомеханические модели предка предков человека и современного человека, то двигательные системы, не имея кардинального анатомического различия, имеют существенные биомеханические различия и особенности.
Модель «Условный общий Предок» — модель «стол» на 4 ногах. Общий центр масс (ОЦМ) располагается непосредственно между 4 опорными конечностями. Для противодействия силе гравитации конечности должны быть разогнуты хотя бы частично. Мышцы-разгибатели конечностей в тонусе, поэтому животное не падает на живот под действием силы тяжести. Нагрузка равномерно распределена между 4 конечностями. В такой модели для 4 конечностей разгибающий [отталкивающий] контур конечностей преобладает над сгибающим. Для туловища наоборот — сгибательный контур доминирует над разгибательным. Туловище не провисает животом вниз по направлению к земле под действием силы тяжести.
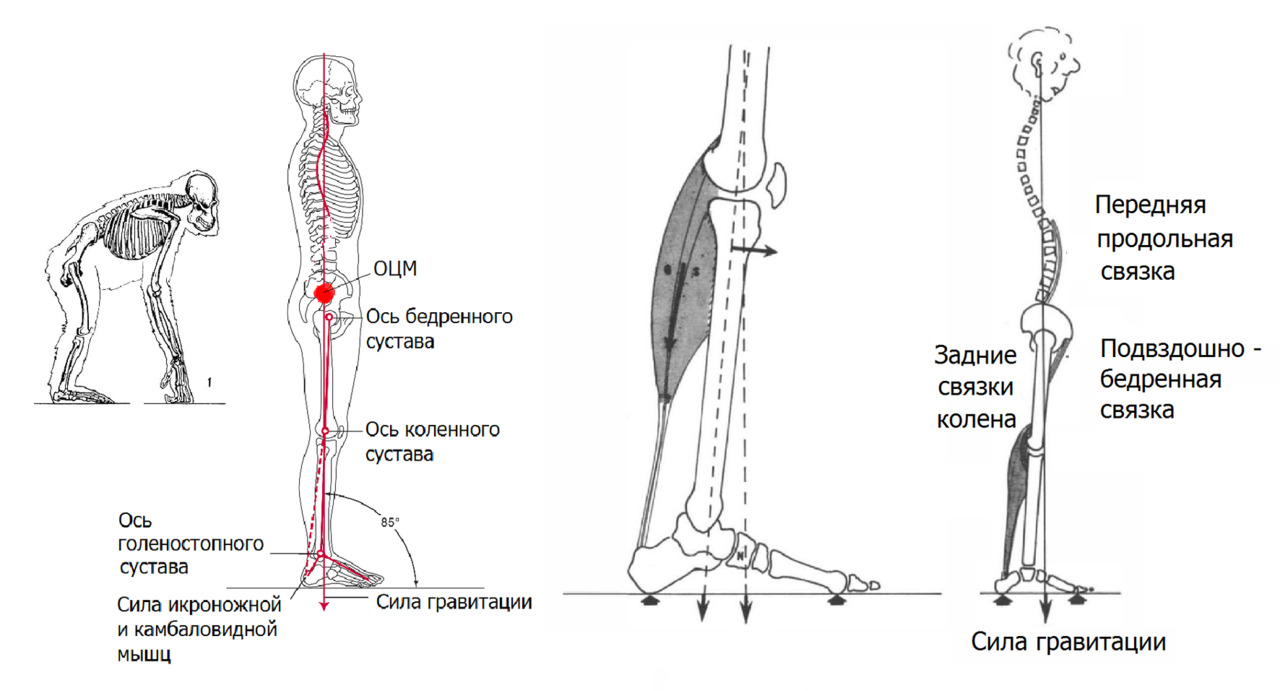
Модель «Условный Человек» — обратный вертикальный маятник, точнее — инерционный многозвенный рычажно-маятниковый механизм с осью вращения ведущих маятников, проходящую через тазобедренные суставы. Верхняя маятниковая система (руки) с осью, проходящую через плечевые суставы, несет вспомогательную стабилизирующую функцию.
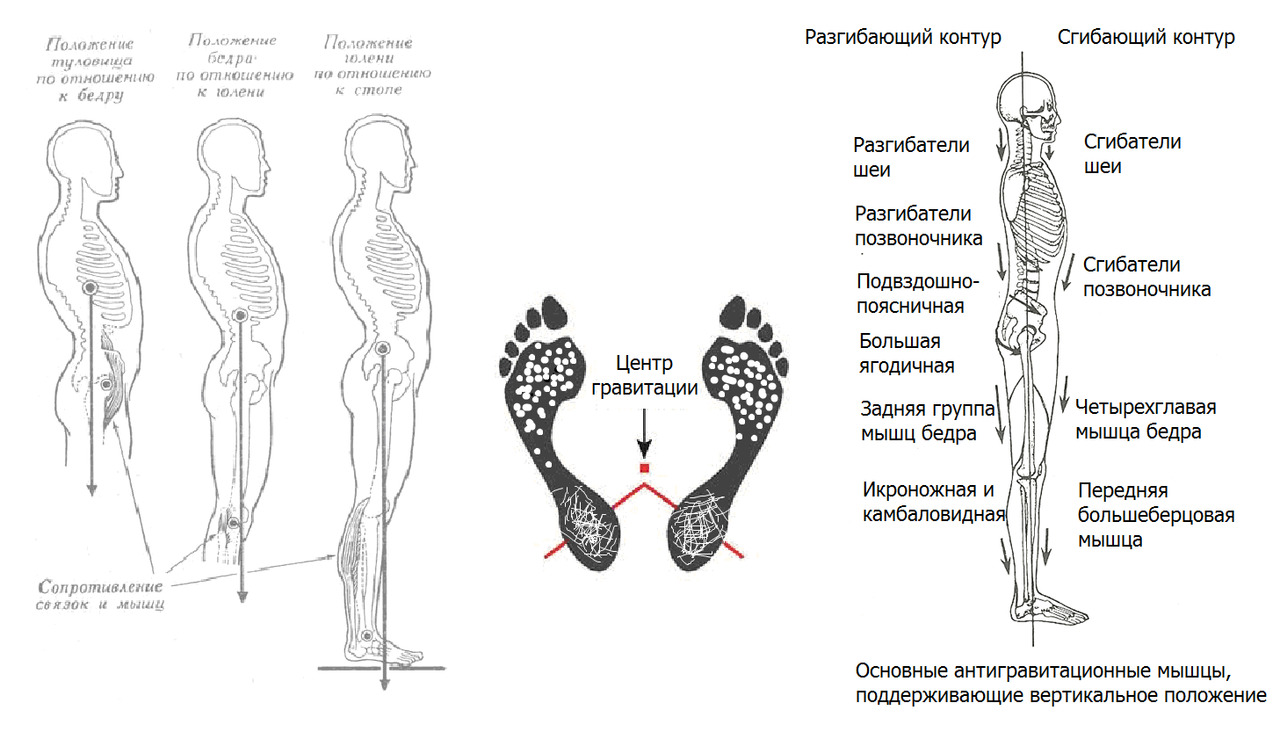
При спокойном положении вертикаль общего центра масс (тяжести) проходит сзади поперечных осей тазобедренных суставов. Голова сбалансирована относительно головок бедренных костей. В расслабленном положении поддержание осанки происходит при минимальном мышечном напряжении. Поддержание вертикальной позы происходит за счет натяжения подвздошно–бедренной связки, подколенных связок и поясничной порции передней продольной связки.
Таз слегка подан вперед, голеностопный сустав замкнут формой таранной кости при легком наклоне вперед.
Вертикально стоящий человек благодаря такой форме позвоночника буквально «висит» на натянутых связках позвоночника и бедра вдоль вектора силы гравитации. Гравитация через натяжение связок «замкнула» позвоночник в положении, для которого не требуется значительная мышечная работа для поддержания вертикального положения.
Коррекция вертикального положения при данной конфигурации осуществляется фоновой тонической работой постуральных мышц, главная из которых камбаловидная мышца голени.
Само по себе поддержание вертикального положения с минимальными затратами не является самоцелью и в чистом виде особой ценности не имеет. Но поддержание оптимального вертикального положения (ортоградного) как части целых двигательный актов имеет решающее значение для локомоции в целом. Качество и экономичность движений зависят от работы постуральной системы, поскольку стабилизация тела считается критическим фактором при выполнении любых задач, особенно динамических.
Любое движение может быть рассмотрено как перемещение из одной равновесной точки (то есть точки, в которой мышечная активность минимальна) в другую.
Выделяют три основных группы двигательных задач:
— задачи, связанные с перемещением базы опоры (ходьба или бег)
— задачи, связанные с неподвижной базой опоры — сидение или стояние
— манипуляционные задачи, требующие выполнения движений верхними конечностями.
Любое перемещение (ходьба или бег) представляет собой переход из нейтрального устойчивого положения в цикл остановленных падений с чередованием фаз опоры и отталкивания с последующим переносом ОЦМ над опорной ногой. Таким образом, ходьба или бег — череда рефлекторно контролируемых «управляемых падений». В начале ходьбы или бега, мышечное сокращение импульсным сокращением смещает ОЦМ в сторону направления движения и выводит тело из равновесного положения — вынуждает тело «падать» в заданную сторону. При каждом шаге человек наклоняется вперед и начинает падение, которому препятствует выдвинутая вперёд нога. После того как она касается земли, на неё переносится вес тела, колено подгибается, амортизируя падение, и выпрямляется, возвращая тело на исходную высоту.
При таком типе локомоции кинетическая энергия движения при постановке ноги рассеивается не полностью, а частично переходит в потенциальную энергию упруго деформирующихся звеньев, которая, в свою очередь, при смене опорной ноги частично преобразуется в кинетическую энергию при отталкивании. Скелет и составляющие его звенья работают как рекуператор энергии, преобразующий часть кинетической энергии движения в потенциальную энергию упругого деформирования скелета и обратно.
(Рекуперация (от лат. Recuperatio) — обратное получение, возвращение энергии, расходуемой при выполнении того или иного технологического процесса, движения, для повторного использования в том же процессе, движении).
При работе в режиме рекуперации, при решении текущей двигательной задачи с целью выйти на заданную величину напряжения включение сократительного аппарата мышцы происходит в режиме сопряжения (наложение фаз во времени) и суммации энергии растянутой соединительной ткани. Мышечное (концентрическое) сокращение синхронизировано с периодом обратного укорочения (возврата к исходной длине) упругих структур мышцы и начинается с началом первой функциональной реакции мышечного аппарата на активацию рефлекса растяжения.
Ортоградное положение и значительный эффект рекуперации энергии определяет механическую эффективность движений человека и выделяет его среди других видов млекопитающих. (К слову, именно эффективность механизма рекуперации энергии препятствует возможности быстро похудеть. Люди потребляют с пищей слишком много калорий, которые просто невозможно израсходовать при низкой двигательной активности).
Сохранение и повторное использование механической энергии происходит за счет действия трех механизмов:
1. Перехода кинетической энергии в потенциальную энергию гравитации и обратно. Экономия энергии в диапазоне 10–25%. Зависит от типа таза и позвоночника, формы и длины костей.
2. Перехода (или передачи) механической энергии от одного звена к другому (напр. от бедра к голени) составляет от 30 до 40% от полной энергии. Зависит от соотношения длины маятников — конечности, длина туловища, масс — инерционные свойства отдельных сегментов.
3. Перехода кинетической энергии движения в потенциальную энергию деформации мышц и сухожилий и обратно. Составляет от 5 до 35%. Зависит от индивидуальной мышечной и связочной композиции. Чем выше процентное соотношение медленно сокращающихся мышечных волокон в составе мышцы, тем выше эффект рекуперации для данной мышцы. Для сухожилий и мышечной оболочки имеет процентное соотношение типов коллагена и эластина.
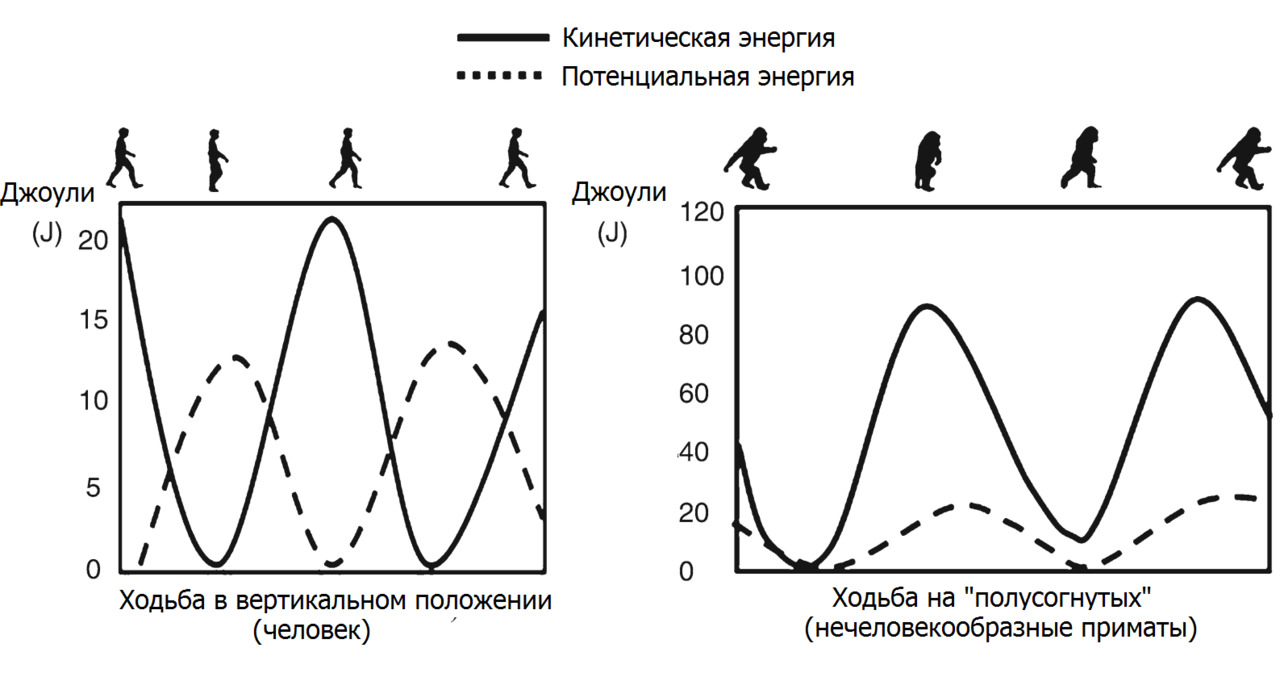
При вертикальном типе передвижения (с оптимальным мышечным балансом) мышечные затраты сведены к минимуму. Скелет человека «запружинен» эластической тягой/натяжением связок, фасций и мышечными соединительно-тканными структурами. Часто такую организацию называют мышечно-фасциальные поезда.
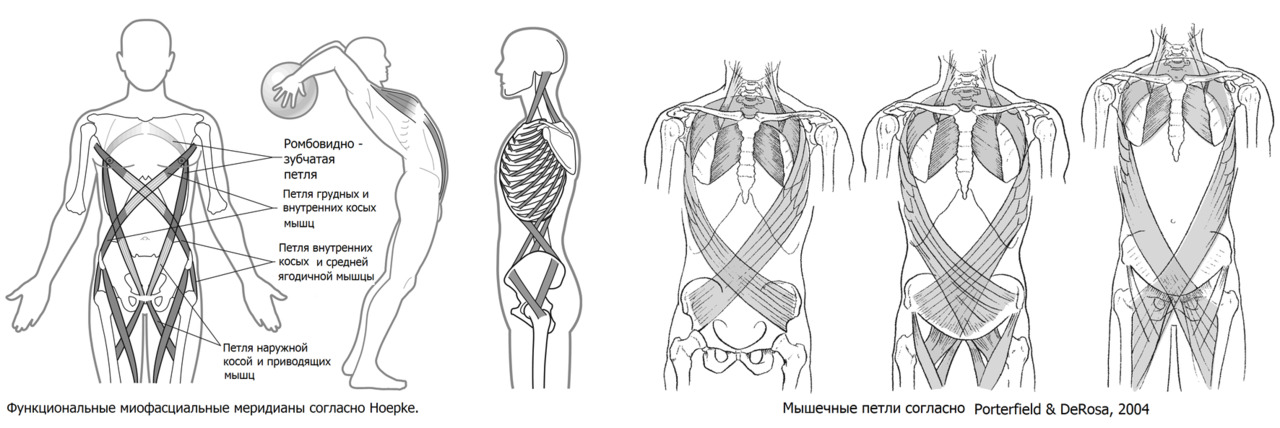
В такой системе происходит передача тягового усилия упруго-эластических компонентов с одних мышечно-фасциальных групп на другие. Такая система построена по типу хиральной системы, в которой все элементы закручены по спирали относительно центральной оси. При движении происходит чередование фаз скручивания-раскручивания с накоплением и отдачей энергии.
Во время бега спортсмен может сохранять около 80% полной механической энергии.
Таким образом, наиболее важным фундаментальным отличием локомоции человека от его эволюционных предшественников является тип передвижения, при котором сила гравитации и реакция опоры (отталкивание от поверхности) преобразуются непосредственно в движение. Система локомоции человека устроена таким образом, что антигравитационная система человека напрямую использует силу гравитации в двигательном акте, преобразуя и контролируя инерцию сегментов тела для осуществления движения с одновременной стабилизацией человека в гравитационном поле.
Эволюционный скачок/рывок представляет собой переход от преимущественно использования мышечной работы с малым КПД в качестве основной движущей силы к использованию условно «бесплатной» силы гравитации с минимальными мышечными затратами на передвижение. Условная «бесплатность» требует пояснения. В этой жизни приходиться платить за все, даже за «бесплатные» вещи.
Чем сложней система, тем более она уязвима к «поломкам» при отсутствии дублирующих механизмов, поддерживающих работоспособность в критических ситуациях. Срабатывает правило: прочность цепи зависит от прочности самого слабого звена. В случае с биокинематической системой человека, самым уязвимым звеном выступает соединительная ткань — связочная и хрящевая. Данный тип ткани относится к брадитрофным, т.е. слабо питающимся и имеющим низкую репаративную способность при повреждении.
Выраженный дисбаланс в регионах скелета, нарушение сопряженности работы в смежных биокинематических звеньях могут привести к мгновенному нарушению в работе отдельных звеньев и выраженному патологическому влиянию на систему в целом.
Те «огрехи», которые «прощаются» гравитацией для локомоторных систем на 4-х конечностях (тип «стол»), не «прощаются» человеку в модели «маятник».
Общепринятая модель перевернутого маятника — сильное упрощение. В такой модели звенья маятника рассматриваются как тонкие твердые тела, вращающиеся вокруг трех идеальных цилиндрических (!) шарниров, моделирующих голеностопный, коленный и тазобедренный суставы. Для математических расчетов вполне подойдет, но в реальности все куда более сложно.
Скелет человека также «сконструирован» с учетом хиральности всех биологических систем — естественной асимметрии, свойства всего живого закручиваться по спирали.
Одно дело, если эта хиральность физиологическая и идет на повышение биокинематичесого энергетического потенциала, другое дело, если «хиральность» искусственного происхождения — «школьные» сколиозы, сколиозирование при отсутствии коррекции выраженной разницы длинны ног и другие механизмы «кручения».
Весь скелет, в той или иной мере, имеет признаки структурной асимметрии. Длинные трубчатые кости у человека тоже имеют винтообразную форму. Это увеличивает прочность и способствует рекуперации. Мышцы вокруг костей закручены по спирали. Позвоночник закручен по часовой или против часовой стрелки. Таз также имеет вид слегка скрученной чаши. Вся система при движении скручивается и раскручивается, накапливая и освобождая энергию. При этом такой механизм препятствует точечному (локальному) износу отдельных структур. Это достигнуто благодаря возможности комбинированных движений. Тазобедренные и плечевые суставы по конструкции — шаровые и обеспечивают три степени свободы. Лучезапястный и голеностопный две степени. Коленный позволяет кручение и скольжение. Подвздошные кости также смещаются относительно крестца во время ходьбы и бега.
Для описания нашей темы подходит другая, более расширенная маятниковая модель. В математических расчетах она не используется, но для раскрытия нашей темы она наиболее уместна.
В силу конструкции позвоночника и суставов, свободно стоящий человек не может остановиться в неподвижном состоянии (нет костного замыкания суставов). Удержание вертикальной позы сопровождается колебательным покачиванием из стороны в сторону относительно вектора гравитации в пределах малых амплитуд. Происходят достаточно сложные взаимовлияющие колебания как общего центра масс (ОЦМ), так и центра давления (ЦД) стоп на плоскость опоры, которые не совпадают по амплитуде и фазности. Это своеобразное равновесное самостабилизирующее раскачивание общего центра масс над площадью опоры. Описывается как «устойчивый конус».
Центр давления (ЦД) — это та интегральная точка на плоскости опоры, в которую объёмное геометрическое тело — человек, имеющий различную плотность тканей организма и постоянно меняющуюся конфигурацию сегментов туловища, как бы «усредняется» в реальном режиме времени в ходе поддержания вертикальной стойки.
Общим свойством ЦД и проекции ОЦМ устойчиво стоящего человека является локализация в пределах координат границы опоры человека (так называемого стопного треугольника).
В условиях постоянно действующего гравитационного поля Земли процесс отклонения тела человека от вертикали в информационном плане является абсолютно необходимым для последующего восстановления утрачиваемого равновесия.
У здорового человека функцию равновесия можно охарактеризовать как устойчивое неравновесие. Основную роль в поддержании «устойчивого неравновесия» выполняет преимущественно тоническая мускулатура. ЦНС «мозаично» перераспределяет напряжение между различными группами постуральных мышц, работающих в аэробном, наиболее экономичном режиме.
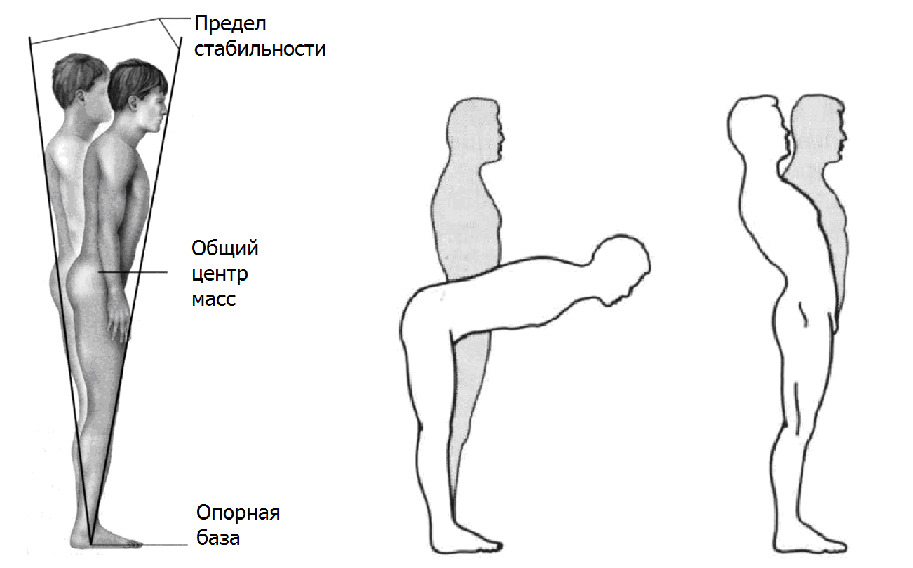
Подобные покачивания незначительны и представляют своеобразную «немую» нейтральную зону, в пределах которой отсутствует активная коррекция положения ОЦМ относительно опоры.
В связи с динамическими взаимодействиями между кинематическими звеньями скелета, отклонение от положения равновесия в любом из отдельно взятых сегментов приводит к необходимости вырабатывать корректирующие силовые моменты во всех суставах. Компенсировать возмущающий фактор и возвращать ОЦМ в равновесную точку.
При увеличении раскачивания или любом произвольном движении, связанном со смещением ОЦМ за площадь опоры, происходит активная коррекция положения ОЦМ или возвратом ОЦМ в проекцию площади опоры за счет мышечного сокращения, или увеличением площади опоры (например, простым расставлением ног более широко).
В вертикальном положение стоящий человек при медленных минимальных возмущениях наиболее часто использует «голеностопную стратегию», рефлекторно (неосознанно) изменяя угол в голеностопном суставе. При быстром возмущении используется «тазобедренная стратегия» с выравниванием через вращение в тазобедренном суставе. Часто используются две стратегии одновременно и автоматически, минуя фокус нашего внимания, т.е. неосознанно.
Колебательный процесс перемещения ЦД осуществляется по плавным дугам, с минимальными затратами энергии, что и характеризует нормальное, комфортное в субъективном плане стояние здорового человека, закрепившего удержание вертикальной позы на уровне прочного автоматизма.
Системы тонической установки тела для обеспечения сложных форм равновесия тела недостаточно. В повседневной и профессиональной (не говоря уже о спортивной) жизни смещение ОЦМ за площадь опоры вещь обыденная и нами редко замечаемая.
Препятствие падению обеспечивается динамическим равновесием или динамической стабилизацией тела в пространстве. Основным отличием её от системы тонической установки тела является возможность реагирования на действие линейных и угловых ускорений — скоростных высокоамплитудных движений в суставах с резким значительным смещением центров масс звеньев тела относительно общего (интегрального) центра масс всего тела.
ЦНС контролирует такое положение посредством тонических рефлексов положения и тонических рефлексов выпрямления.
Поддержание равновесия — удержание туловища против действия силы тяжести и способность контролировать и стабилизировать положение различных частей тела относительно друг друга во время сложных движений достигается за счет координированной работы мышечной и вестибулярной системы. Зрительная система является неспецифической для данного вида деятельности и выступает как фон.
Для поддержания равновесия тела одну из решающих ролей играет информация о гравитационной вертикали от вестибулярного аппарата и проприоцепторов тела при контакте с плоскостью опоры. Для формирования чувства равновесия необходима согласованность информации от полукружных каналов ушного лабиринта, слуховых рецепторов, проприоцепторов мышц шеи, механорецепторов связок, сухожилий и сетчатки глаз.
В результате суммирования тонических отолитовых и шейных рефлексов положения всякой позиции головы в пространстве и по отношению к туловищу соответствует совершенно определённое перераспределение тонуса мышц конечностей и глаз.
Проприоцепторы передают информацию о позе тела в пространстве и расположение конечностей относительно туловища (чувство положения). Кроме того, они регистрируют положения в суставах и их скорость (чувство движения) и могут оценивать приложенное мышечное усилие (чувство силы). Эта информация далее направляется вместе с импульсами от органов чувств и равновесия в высшие центры моторной деятельности (мозжечок и т.д.). Эти импульсы передают основную информацию для контроля движений, коррекции поддержания позы и целенаправленных действий.
Взаимодействие рефлексов положения и выпрямления обеспечивает оптимальную ригидность (жесткость) тела и закономерное перераспределение мышечного тонуса тела, конечностей и глаз в зависимости от положения относительно вектора гравитации, ощущения плоскости опоры и зрительной фиксации окружающих предметов.
Поддержание вертикальной позы — это двигательная функция, представляющая собой статическое движение, обеспечиваемое фоновой тонической мышечной активностью преимущественно разгибателей, которые за счет непрерывного взаимного перемещения звеньев тела поддерживают ОЦМ относительно площади опоры.
Если же постуральная система человека функционирует негармонично/не слаженно либо имеет место тот или иной патологический процесс, поразивший структуры, обеспечивающие функции равновесия, то в поддержании вертикальной позы дополнительно задействуется и фазическая мускулатура, в норме задействованная только при динамической стабилизации. Это приводит к снижению точности контроля выполнения движений и требует не только гораздо большего расхода энергии, но, при этом, увеличивая общую компрессионную нагрузку на опорные структуры скелета.
Вполне логичный вопрос: какое отношение это имеет к болям в спине и остеохондрозу в частности? Самое непосредственное. И не только к остеохондрозу, но и к другим поражениям опорно-двигательного аппарата: коксартрозам, гонартрозам, плоскостопию, сколиозированию и пр. Чем выше доля участия сократительного аппарата мышцы в стабилизации сустава или поддержании вертикальной позы, тем выше компрессионная нагрузка на суставы и межпозвонковые диски. Длительная компрессионная нагрузка приводит к резкому ухудшению питания и трофики в мягких тканях и запуску дегенеративно-дистрофических процессов. Происходит необратимый сдвиг обменных процессов в сторону катаболизма и необратимой деградации связочной и хрящевой ткани.
ГЛАВА 6. МЕХАНИЗМЫ РАЗВИТИЯ «НЕСПЕЦИФИЧЕСКОЙ» БОЛИ В СПИНЕ
Мне очень часто приходится слышать жалобы пациентов на то, что врачи толком не могут объяснить причины боли или, что лечение безрезультатно. Также многие пациенты выражают недовольство тем, что при смене врача новый врач начинает опровергать объяснения предыдущего и дает объяснение по-своему. Поскольку лечение может затянуться, то при попытках выяснить причины хронизации боли пациенты начинают самостоятельно выискивать информацию в интернете или консультируются с такими специалистами как массажисты, тренера фитнесс-центров, мануальщики и пр., что еще больше запутывает пациентов и повергает в уныние и депрессию.
Все это бесперспективно и не имеет никакого смысла.
Как я уже говорил раньше, все лечение боли в спине пытаются переложить на плечи семейных врачей и терапевтов. В крайних случаях (при выявлении серьезной патологии) пациенты будут перенаправляться к неврологам или другим смежным специалистам. Одной из причин, по которой лечение болей в спине «поручают» семейным врачам и терапевтам, является их низкий уровень знаний в области вертеброневрологии. Когда врач не знает и не понимает причин и механизмов боли и патологии позвоночника, то это как раз именно тот специалист, который строго будет придерживаться установленных алгоритмов лечения, и не будет «своевольничать» и подстраивать схемы под каждого конкретного пациента. Что, собственно, и нужно заинтересованным лицам. Перед данными специалистами не стоит задача вылечить. Главная их задача — выписать нужный препарат. Второй причиной, по которому никто из вышеупомянутых специалистов не сможет что-либо внятно объяснить является то, что все отечественное медицинское терапевтическое мировоззрение строится на физиологии Павлова и примитивном учении о рефлекторных дугах. Опираясь на устаревшее учение о рефлексах объяснить причины боли рядовому семейному врачу очень сложно (попросту невозможно), но для выписки рецепта устаревшей концепции вполне хватит.
Для вертеброневрологии как отрасли медицины физиология Павлова не подходит, — она слишком примитивно объясняет все процессы. Для объяснения многих болевых и неврологических феноменов подходит физиология движения Н. А. Бернштейна и теория функциональных систем П. А. Анохина. Хронизация боли — работы А. Р. Лурия, С. Н. Крыжановского, И. М. Фейгенберга, О. С. Андрианова, Р. Мелзака.
Поэтому И. Р. Шмидт (2004) делает такой вывод:
«При изучении роли центра и периферии в реализации остеохондроза позвоночника (ОП), неврологических проявлений (ОП) и сопровождающих их патобиомеханических изменений, было установлено, что врожденный дефицит преднастройки и вероятностного прогнозирования, моторная неодаренность, функциональная асимметрия мозга, особенности личности и личностного реагирования, а также искаженная проприоцептивная афферентация оказывают существенное влияние на реализацию генетической предрасположенности к ОП и на клинический полиморфизм неврологических проявлений ОП. В формировании неврологических и патобиомеханических синдромов участвуют все пять уровней организации движений, по Н. А. Бернштейну».
Я думаю, что после изучения главы читатель поймет, почему не дали развиться такой специальности как «вертеброневрология» и почему всячески льют грязь как на вертеброневрологов, так и на Я. Ю. Попелянского в частности.
Поскольку читатель, вероятнее всего, о физиологии движения, разработанной выдающимся советским физиологом Николаем Александровичем Бернштейном, ничего не слышал, то я кратко представлю учение об уровнях организации движения.
Уровни организации движения по Н. А. Бернштейну (1947).
А — Уровень тонуса (руброспинальный) — организация движений в поле земного тяготения.
Уровень определяет функциональные возможности мускулатуры через преднастройку мышечного тонуса, влияя на сегментарный аппарат спинного мозга и фоновое изменение его возбудимости.
Отвечает за реципрокную (взаимозависимую, взаимовлияющую) иннервацию мышц, регулирует силовые и скоростные характеристики сокращения мышц. Абсолютный монополист по тонусу мышц во всей центральной нервной системе.
B — Уровень синергий (усиление эффекта за счет взаимовлияния; содружественное и взаимодополняющее действие (таламо-паллидарный)) — организует движения в системе координат собственного тела, частей тела относительно друг друга посредством обширных мышечных синергий, обеспечивающих согласованную работу многих десятков мышц.
Преодолевает и использует реактивные силы в управлении многозвенными маятниками конечностей.
Использует стойкие штампы и автоматизмы, контролирует динамическую устойчивость движений, которые требуют точной согласованности ритмически повторяющихся во времени сложных комбинаций движений, охватывающих все конечности и туловище (ходьба, бег, плавание и другие).
Точно воспроизводит один и тот же пространственный и ритмический рисунок движения.
Способен к согласованию мышечных усилий в опорном положении, вне зависимости от информации, поступающей от телерецепторов (глаза, слух).
Контролирует всю внутреннюю черновую технику сложных движений, определяя индивидуальные особенности моторики.
C — Уровень пространственного поля (пирамидно-стриальный) — организовывает движения в системе координат окружающего пространства: координат, связывающих тело и его части с предметами пространства и сами предметы между собой.
Отвечает за организацию движений частей тела или всего тела относительно предметов пространства, точность и меткость в овладении пространством, заполненным объектами, имеющими размер, форму и массу. Движения уровня пространственного поля имеют ясно выраженный целевой характер, вектор, координату в пространстве.
Использует дистантные рецепторы (глаза).
D — Уровень действий (теменно-премоторный) — организует движения собственного тела и его частей с целью приведения в движение окружающих предметов относительно друг друга в соответствии со смысловой задачей, смысловой сущностью предметов, их свойствами и объективно закономерными внутренними и внешними связями. Уровень манипуляции с предметом — «ручное» мышление.
Е — Абстрактный уровень — символические координации — организация движений в соответствии с мыслимой целью движения, воспроизведение образов предметов и явлений посредством речи, письма.
За выполнением любой двигательной задачи отвечает моторный контроль. Высшие мозговые центры выполняют управляющую роль относительно нижерасположенных центров. Любая задача решается через выполнение моторных программ, которые централизованно хранятся внутри многочисленных моторных областей мозга и частях ЦНС.
Моторные области развиваются у человека от нижележащих отделов (спинной мозг) к вышележащим (ассоциативные области коры головного мозга) и деградируют в обратном порядке от вышележащих к нижележащим.
Для построения движений различной сложности команды отдаются на различных уровнях нервной системы. Выполнение моторной активности инициируется, координируется и контролируется центральными механизмами, которые действуют на основе иерархии. Каждый уровень выполняет свою строго специфическую функцию; при этом он может воздействовать как на низшие, так и на высшие уровни в зависимости от поставленной двигательной задачи, выступая управляющим уровнем.
Структуры ЦНС на протяжении жизни накапливают необходимые модели/шаблоны двигательных актов, формируют и хранят «заготовки» программ по реализации различных двигательных стратегий.
Привычно реализуемые шаблоны воспроизводятся как стереотипы. Статичных стереотипов, установленных по умолчанию, не существует.
Любое целенаправленное действие представляет собой последовательное выполнение моторных программ, состоящих из шаблонов с коррекцией на текущую ситуацию.
Когда наступает момент выполнить какое-либо действие, основной шаблон или список моторных заготовок уже существует в ЦНС. В связи с требуемыми обстоятельствами он модифицируется по силе, времени и направлению, что приводит к выполнению уже нового движения, но по выверенным правилам для выполнения данной группы задач. Вариант импровизации. Повторение без повторения на основе имеющихся заготовок.
Моторные программы условно делятся на генерализованные — локомоторные и задача-специфические — разовые неповторяемые.
Моторные программы в ЦНС не содержат специфических движений, но включают общие правила для специфического класса движений. Наиболее часто используемые программы выполняются наиболее координировано и с наименьшей энергетической стоимостью и наименьшими погрешностями.
Вокруг значимой двигательной задачи происходит динамическая самоорганизация функциональных систем и субсистем, обеспечивающих выполнение целенаправленного действия для решения определенной двигательной задачи. Для реализации движения все системы должны работать синхронно и при этом не истощать смежные системы.
Для согласованной работы всех уровней организации движения ЦНС должна оперировать точными данными с периферии со всех подконтрольных структур.
Сенсорный (афферентный) поток непрерывно идет во все анализирующие центры головного мозга.
Для точного управления движением ЦНС непрерывно анализирует параллельные потоки сенсорной информации, идущей от связочного аппарата, мышц, кожных покровов, вестибулярного аппарата и глаз.
Сенсорная информация, идущая из связок, сухожилий, капсул суставов, фасций и межпозвонковых дисков имеет первостепенное значение в планировании будущего движения. На основе данной информации ЦНС выбирает из списка имеющихся шаблонов группу шаблонов, на основе которых будет «сконструировано» и реализовано будущее движение. Можно сказать, что информация с сухожильно-связочного аппарата используется для прогнозирования будущих двигательных сценариев, планирования движения и подготовки упреждающих команд.
Информация, поступающая с мышц, используется для коррекции движения непосредственно при выполнении самого движения.
Информация с кожных рецепторов является уточняющей. Разделение сенсорных потоков условно и применено для облегчения восприятия. ЦНС рассматривает всю входящую информацию цельным двигательным сенсорным паттерном («рисунком»).
Благодаря совершенству моторного контроля человек выполняет сложные двигательные задачи с минимальными энергозатратами и с минимальным износом опорных и миофасциальных структур скелета, но при условии, что все системы и подсистемы работают слаженно и в оптимальном режиме.
Введение
Механизмы развития «простой» неспецифической боли я разобью на отдельные составляющие/механизмы для удобства восприятия. Очень часто они встречаются одновременно, усиливая эффект и затрудняя выявление ведущего.
Основным виновником неспецифической боли являются мышцы, связки и капсулы суставов — ткани, имеющие наибольшую иннервацию свободными нервными «болевыми» окончаниями. Принято считать, что чем обильней иннервирована ткань, тем больший вклад в развитие болевых феноменов она вносит. Для позвоночника в этом плане первое место занимают капсулы суставов, второе — задняя продольная связка и диск, и третье — менискоиды — прокладки внутри сустава по типу менисков в коленном суставе. Обильная иннервация плотной оформленной соединительной ткани (фиброзной) не случайна. Данный тип ткани относится к брадитрофным, т.е. таким, для которых характерен низкий уровень питания. Соответственно, при повреждении — очень низкая репаративная способность. Поврежденный сухожильный и связочный аппарат может не восстанавливаться годами. Обильная иннервация [выполняющая сигнальную функцию], таким образом, защищает соединительную [хрящевую] ткань от возможного регулярного повреждения. Незначительная травма приводит к выраженной боли и обездвиживанию (шинированию) поврежденного участка. Другого способа защиты соединительной ткани у организма нет.
С мышцами в этом плане проще: они хоть и восстанавливаются с некоторым включением фиброзной ткани, но выраженной потери функции (при правильном лечении), как правило, не наблюдается. Вовремя не устраненное нарушение в работе мышц приводит к травматизации хрящевых и капсульных структур, накоплению участков атрофий и зон износа связочного аппарата. Данные патологические изменения приводят к снижению функциональных возможностей звеньев скелета, что может значительно влиять на биомеханику позвоночника.
Наиболее удобно позвоночник рассматривать через разделение на субсистемы, обеспечивающие основные функции позвоночника. Разделение условно и используется только для упрощения восприятия.
Концепция Панджаби (Manohar Panjabi). Стабилизирующие системы позвоночника.
Стабилизация тела и позвоночника в пространстве относительно вектора гравитации зависит от слаженной работы трех взаимосвязанных и взаимозависимых субсистем.
Пассивная субсистема включает в себя позвонки, межпозвонковые суставы, межпозвонковые диски, связочный аппарат позвоночника, капсулы суставов. Также к пассивной части относят глубокие и поверхностные фасции и мышечные соединительнотканные оболочки.
Активная скелетно-мышечная субсистема состоит из мышц и сухожилий, прикрепляющихся как непосредственно к позвоночнику, так и к другим костным рычагам опорно-двигательной системы.
Невральная субсистема или субсистема обратной связи состоит из рецепторов, информирующих ЦНС о параметрах силы, угловых ускорениях, силе и скорости растяжения/сокращения мышц и многое другое. Данные рецепторы располагаются в мышцах, связках, сухожилиях, капсулах суставов, фасциях.
Все три субсистемы работают синхронно, обеспечивая статическую и динамическую стабилизацию тела в пространстве и позвоночника в частности.
Поскольку человек постоянно находится в состоянии контролируемого неравновесия, то для каждого сустава и межпозвонкового диска существует точка (центр реакции), где суммируются моменты сил гравитации, мышечной тяги, груза и т. д. Невральная контролирующая субсистема отслеживает все параметры с механорецепторов, расположенных в связочной ткани и проприоцепторов, расположенных в мышечной ткани, с целью равномерного распределения нагрузки на отдельные суставы и максимально их минимизирует.
Всегда нужно помнить, что конструкция «модель человека — маятник» прежде всего костно-связочно-фасциальная, в которой сократительный компонент мышц играет второстепенную роль исполнителя или корректора движений. Только часть движений выполняется за счет активации сократительного компонента мышц, в то время как другая часть выполняется как за счет внешних сил (гравитация, инерция), так и внутренних (упруго-эластические свойства соединительной ткани). Иногда мышцы только начинают движение, а гравитация и инерционные силы его выполняют. Мышцы в данной модели — модулятор и корректор положения и движения. Избыточная мышечная активация приводит к увеличению нагрузки на суставы, что может привести к быстрому износу опорных структур — дисков, менисков и гиалиновых хрящей суставов.
Для удобства объяснения причин боли, механизмы я разобью на три части, согласно стабилизирующим системам позвоночника: боль мышечная, фасциально-связочно-дисковая, и боль при поражении невральных структур.
Сначала стоит рассмотреть механизм развития неспецифической боли, вызванной нарушением мышечной работы
Чтобы понять причину мышечной боли, следует бегло вспомнить некоторые необходимые факты о мышцах:
Для нашей темы достаточно рассмотреть мышцы через 3-х компонентную усредненную модель.
Любая мышца состоит из сократимой, эластичной и сухожильной ткани.
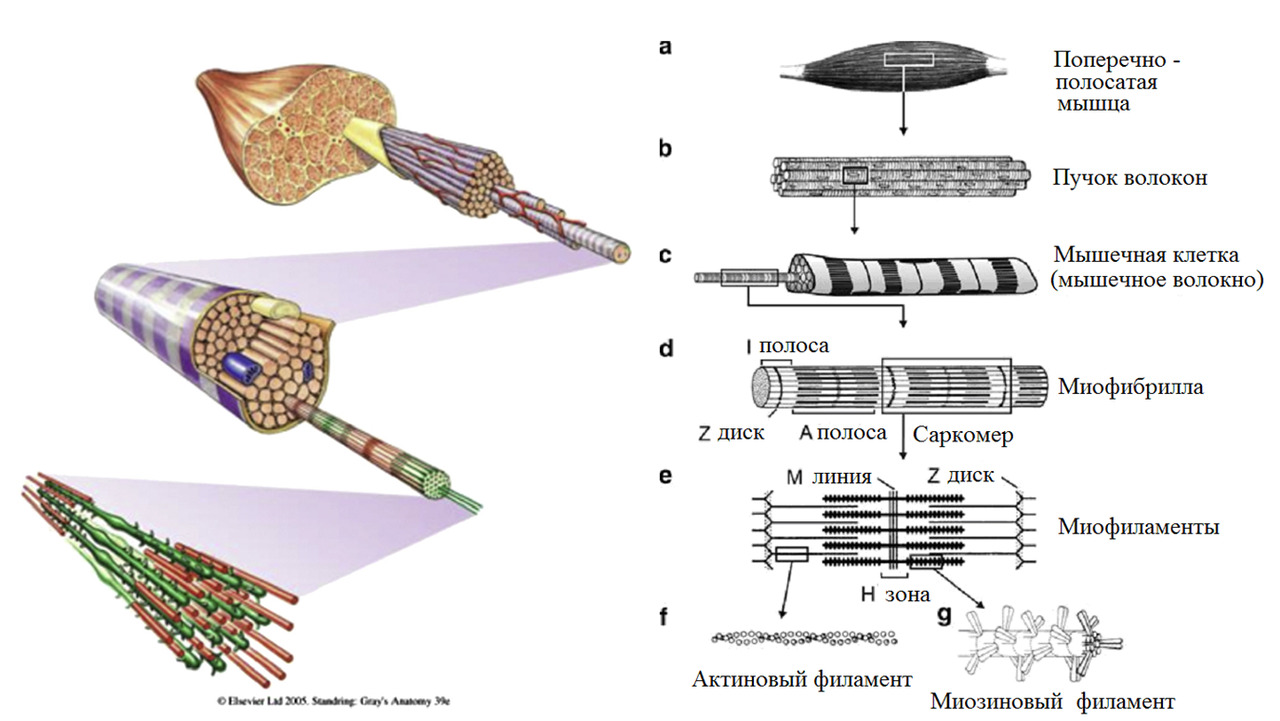
Мышечные сократительные волокна (мышечные клетки) подразделяют на два типа: экстрафузальные и интрафузальные.
Экстрафузальные — основной сократительный субстрат, вызывающий сокращение мышцы. Т.е. тот компонент, который обывателем воспринимается непосредственно как «мышца» или «мясо».

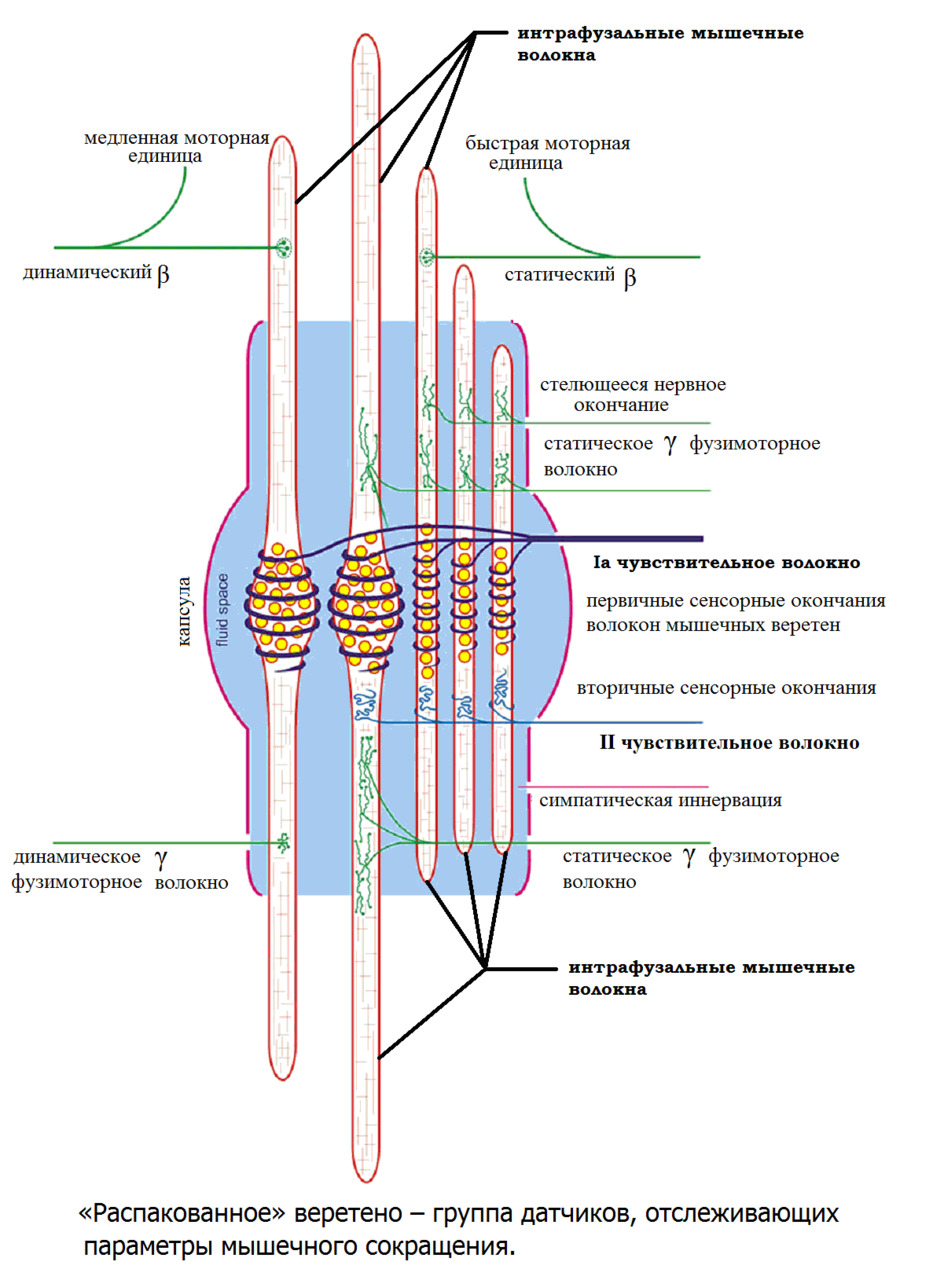
Интрафузальные (внутри веретена) — тонкие мышечные волокна, через напряжение/расслабление которых происходит настройка и регулировка чувствительности датчиков длины и скорости сокращения мышцы. Так называемое «веретено». Вклад в развитие силы вносят опосредовано, участвуя в реализации рефлекса растяжения (миотатический рефлекс).
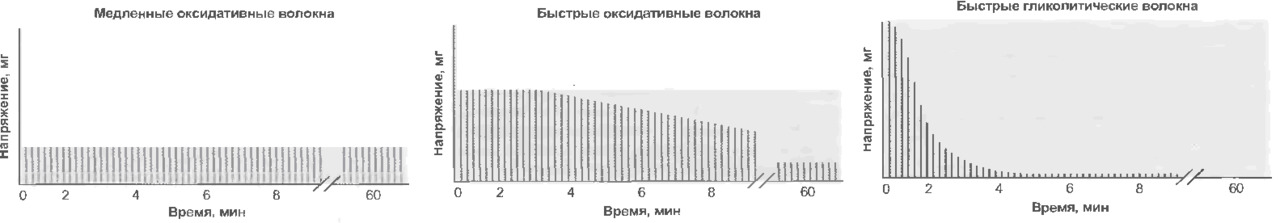
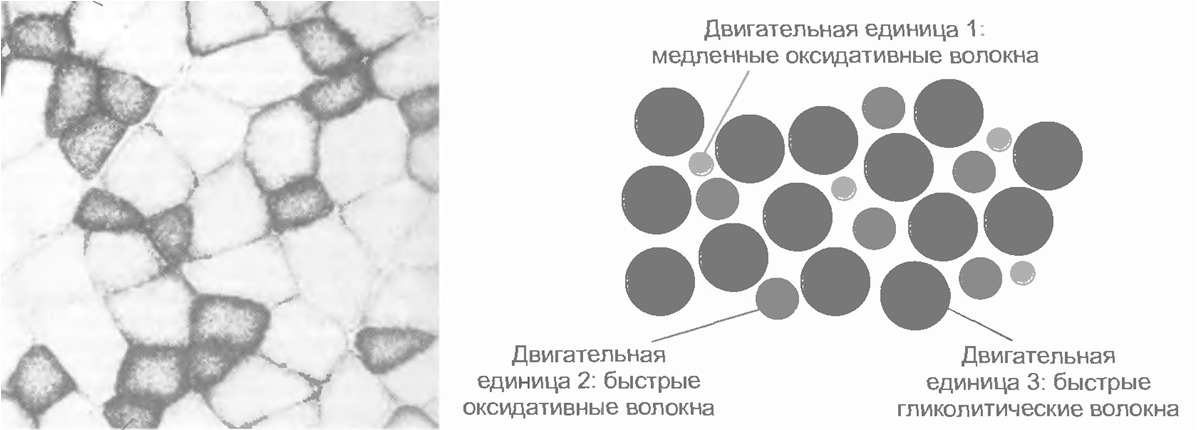
Экстрафузальные мышечные волокна подразделяются на три типа с разными характеристиками.
1. Медленные оксидативные волокна (I). Волокна I типа генерируют самую малую силу, но имеют наибольшую выносливость. Обеспечивают длительное сокращение мышцы; это используется для поддержания позы. Мембрана волокна не обладает электрической возбудимостью. Активация зависит от частоты стимуляции. Для данного типа волокна характерно медленное градуальное сокращение и медленное расслабление (приблизительно в 100 раз медленнее, чем у быстрых волокон).
Тип энергообеспечения — аэробное дыхание. Многие волокна при недостатке О2 продолжают работать за счет анаэробного гликолиза; в этом случае образуется лактат (молочная кислота) и создается кислородная задолженность.
2. Быстрые гликолитические волокна (IIb) Волокна II b обеспечивают самую большую силу, но обладают наименьшей выносливостью. Обеспечивают немедленное быстрое сокращение. Незаменимы при динамических нагрузках и скоростной локомоции. Мембрана волокна обладает электрической возбудимостью. Когда возникает активация волокна, развивается ответ типа «все или ничего». Мгновенное сокращение всех волокон в моторной единице. В качестве источника энергии интенсивно используется гликоген (анаэробный гликолиз, образуется лактат (молочная кислота). Сокращение в три раза быстрее, чем у медленных волокон. Довольно быстрое утомление.
3. Быстрые оксидативные волокна (IIa) Волокна IIa тип — универсальный. Обладают характеристиками как 1 так и 2 типа.
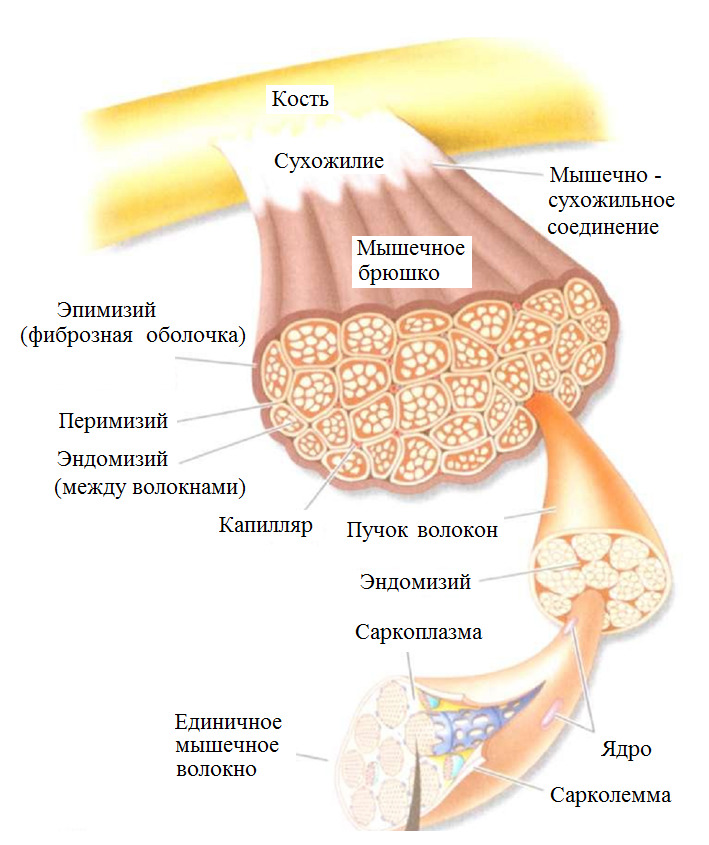
Каждое мышечное волокно окружено соединительной тканью (эндомизиумом). Пучки волокон также покрыты соединительно-тканной оболочкой (перимизиумом). Пучки объединены в целую мышцу с помощью эпимизиума. Вся мышца покрыта фасцией. Общая форма мышцы определяется соединительной тканью. Соединительная ткань преобладает над мышечными волокнами.
Эластические элементы подразделяют на последовательные и параллельные элементы. Последовательные эластические элементы (сухожилия) действуют по одной оси с сократительными элементами.
Параллельные эластические элементы (эндомизиум, перимизиум и эпимизиум) действуют параллельно оси действия сократительных элементов.
Каждая мышца состоит из трех типов мышечных волокон в разной пропорции. Зависит от глубины залегания и выполняемой функции. Чем «глубже» расположена мышца, тем больше в ней содержание медленных волокон. В поверхностных мышцах больше быстрых волокон. Мышечная композиция генетически предопределена.
Сила, которую мышца может развить благодаря своим эластическим элементам, будет максимальной, если мышца предварительно эксцентрически растянута внешней силой.
Сила, которую мышца может развить благодаря своим сократительным элементам, будет наибольшей, когда мышца начинает сокращение при длине, соответствующей состоянию покоя.
Общая сила, развиваемая совместно как эластическими, так и сократительными элементами, будет максимальной, если мышца предварительно эксцентрически растянута.
Эксцентрическая работа является работой сопротивления мышцы к ее растяжению внешней силой.
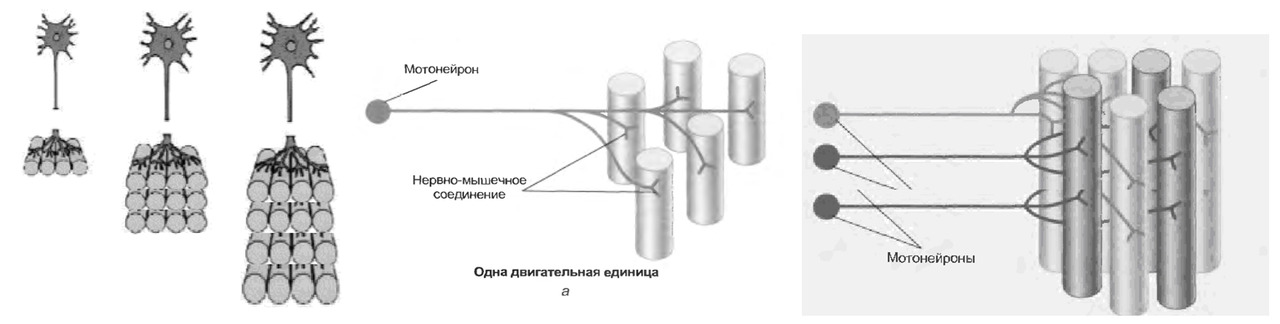
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов — нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления — аксоны в составе двигательного нерва подходят к мышце.
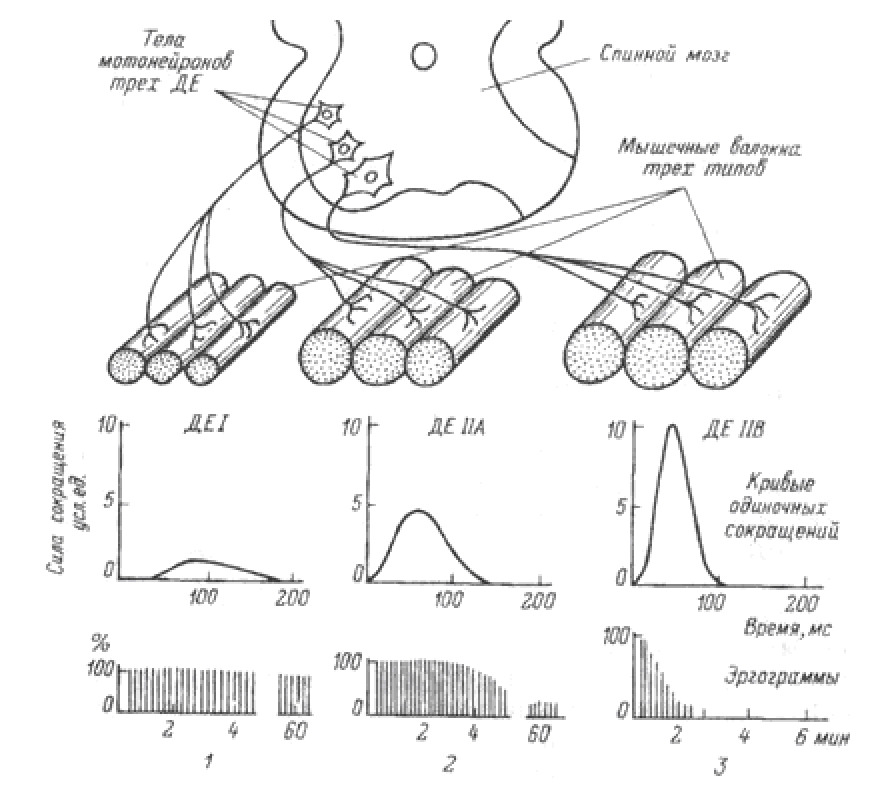
Система, включающая в себя альфа-мотонейрон, аксон и совокупность мышечных волокон, которые он иннервирует, получила название Моторная единица (МЕ) или двигательная единица (ДЕ).
Войдя в мышцу, аксон моторного нейрона разветвляется на множество веточек, каждая из которых подведена к отдельному волокну. Таким образом, один мотонейрон иннервирует целую группу волокон, которые сокращаются/расслабляются как единое целое.
Различают три типа моторных единиц.
Малый мотонейрон с малым количеством мышечных волокон. Имеет высокую степень возбудимости (низкий порог активации). Иннервирует медленные оксидативные волокна (кислород зависимые) (тип I).
Средний мотонейрон со средним количеством мышечных волокон. Имеет промежуточную степень возбудимости (средний порог активации). Иннервирует быстрые оксидативные волокна (тип II a).
Большой мотонейрон с большим количеством мышечных волокон. Имеет низкую возбудимость (высокий порог активации). Иннервирует быстрые гликолитические волокна (кислород независимые) (тип IIb).
Мышечные волокна отдельной моторной единицы всегда одного типа.
Мышца сокращается или расслабляется при взаимодействии двигательных и чувствительных нейронов.
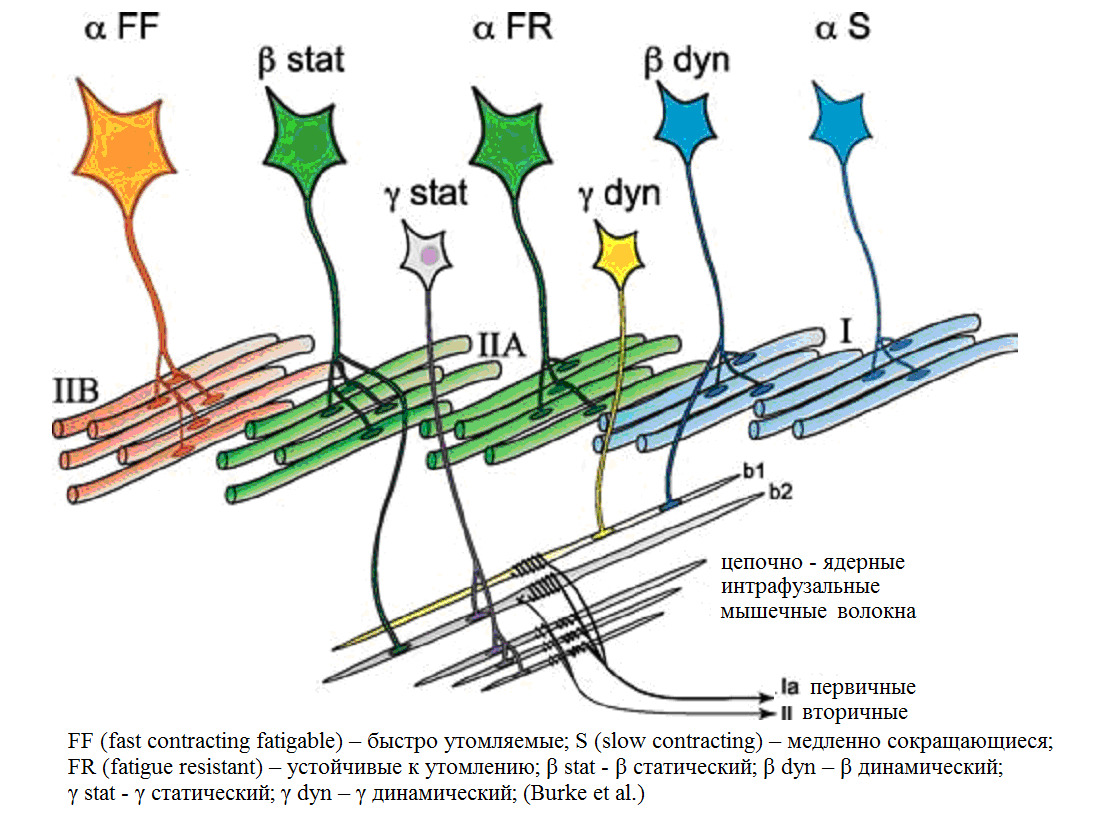
(Manuel, M., & Zytnicki, D. (2011). Alpha, beta and gamma motoneurons: functional diversity in the motor system’s final pathway. Journal of integrative neuroscience, 10 (03), 243—276).
Альфа (α) -мотонейроны иннервируют экстрафузальные волокна. Гамма (γ) -мотонейроны — иннервируют интрафузальные. Бета (β) — одновременно иннервируют как экстра, — так и интрафузальные. Отвечают за слаженность сокращения обоих типов волокон, предотвращают потерю чувствительности «веретена» при сокращении экстрафузальных.
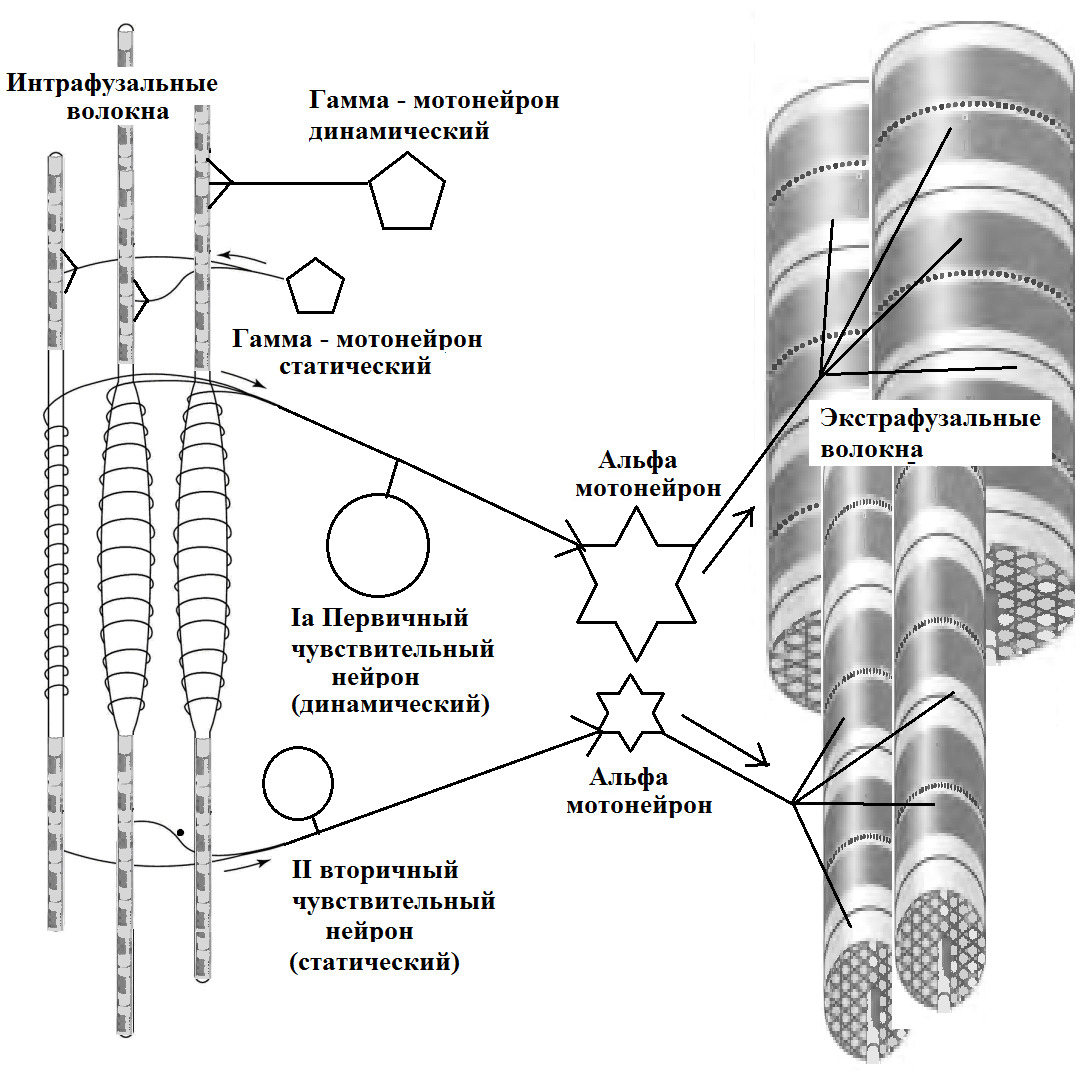
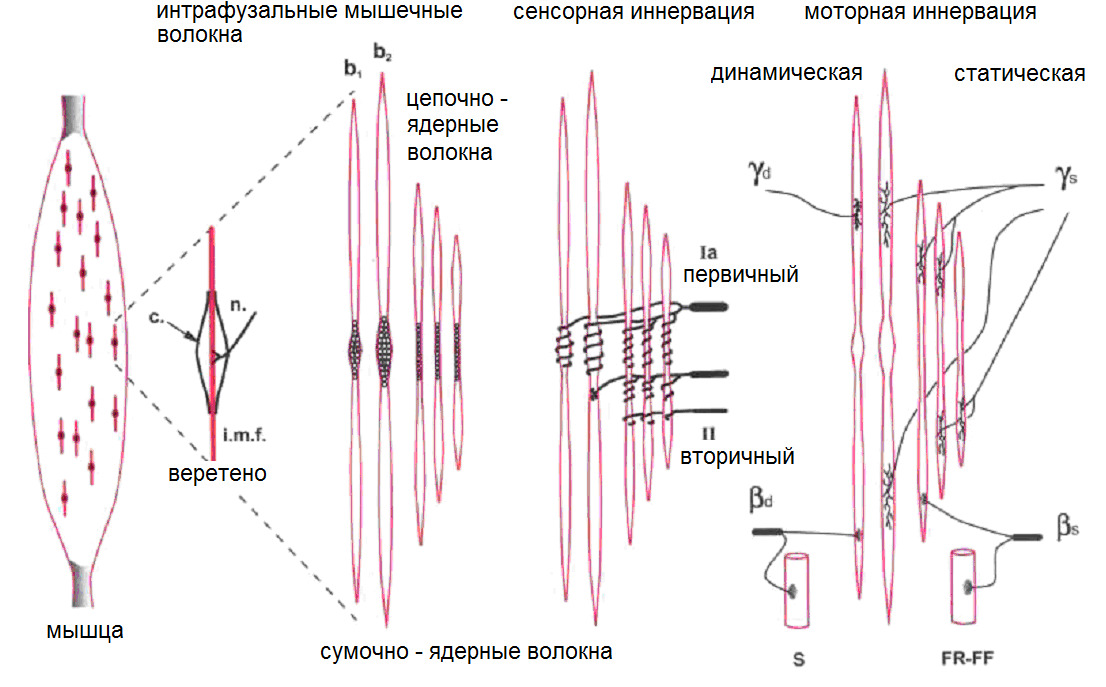
(Ellaway, P. H., Taylor, A., & Durbaba, R. (2015). Muscle spindle and fusimotor activity in locomotion. Journal of anatomy, 227 (2), 157—166).
В норме для развития максимальной силы мышцы должны активироваться все моторные единицы.
Функциональные свойства разных моторных единиц отражают особенности составляющих их мышечных волокон.
Совокупность мотонейронов, связанных с одной мышцей, называется пул мотонейронов. В нем представлены все три типа моторных единиц.
Синхронизация. При возбуждении одного альфа мотонейрона все иннервируемые им мышечные волокна одновременно сокращаются. Сила сокращения всех этих волокон будет суммироваться.
Последовательность активации моторных единиц называется рекрутинг.
Первыми активируются самые малые альфа-мотонейроны, связанные с медленными оксидативными волокнами.
Затем включаются быстрые оксидативные, чьи волокна крупнее. И наконец, активируются самые крупные мотонейроны, иннервирующие быстрые гликолитические волокна.
Сокращение медленных волокон может развивать до 15% максимальной силы мышцы.
Сокращение быстрых оксидативных может развивать до 45% максимальной силы.
Сокращение быстрых гликолитических волокон может обеспечивать максимальную силу сокращения.
Управлять силой мышечного сокращения можно только через усиление или ослабление частоты нервного импульса, вовлекая в работу новые моторные (двигательные) единицы, либо наоборот, выключая из работы задействованные МЕ.
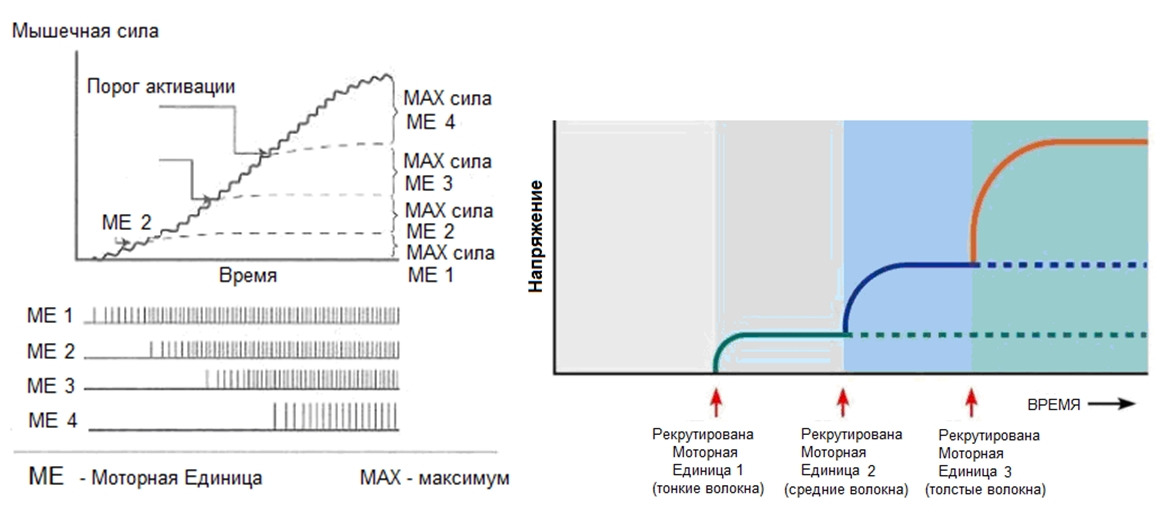
Каждая последующая по уровню порога активации моторная единица имеет большее количество мышечных волокон. Причем разница по количеству мышечных волокон между соседними моторными единицами растёт с увеличением порога возбудимости (активации) и если низкопроговые моторные единицы могут разниться в несколько мышечных волокон, то высокопороговые моторные единицы разнятся между собой уже в десятки или сотни мышечных волокон. Соответственно, активация таких волокон уже будет иметь не «дискретный» (градуированный) характер, а «взрывной».
У нетренированных людей количество одновременно сокращенных волокон не превышает 17%. В результате физических тренировок у высококвалифицированных спортсменов количество одномоментно активированных волокон составляет порядка 40%.
Исходя из этого можно сделать вывод, что наиболее точные и координированные движения можно совершать при наименьшем мышечном напряжении, когда, рекрутируя новые моторные единицы, ЦНС регулирует силу мышцы десятками новых мышечных волокон, вовлеченных в работу, а не тысячами.
Нарушение механизма последовательной активации/деактивации мышечных волокон серьезно влияет на всю систему управления движением.
Количество мышечных волокон, принадлежащих одной и той же моторной единице, увеличивается с возрастом. Это обусловлено изменением коэффициента иннервации, поскольку некоторые двигательные нейроны вырождаются и умирают, что приводит к тому, что аксоны других моторных единиц образуют большие ответвления и реиннервируют мышечные волокна, утратившие аксональную связь (происходит разрастание аксонов с захватом «чужих» мышечных волокон). Таким образом, с возрастом количество моторных единиц уменьшается, тогда как размер (количество мышечных волокон) тех моторных единиц, которые остаются, — увеличивается.
При постуральной (позной) деятельности в работу вовлекаются преимущественно низкопороговые медленные энергоэкономичные устойчивые к утомлению моторные единицы, а мышечные волокна, входящие в состав таких единиц, функционируют в режиме, близком к изометрическому и развивают при этом длительные тетанические сокращения небольшой силы, т.е. находятся в режиме постоянного сокращения.
В мышцах, для функции которых необходимы кратковременные сильные сокращения, больше быстрых волокон (фазные мышцы).
Рекрутинг — механизм последовательного включения сократительного аппарата мышцы, напрямую зависящий от выполнения поставленной на текущий период двигательной задачи (выдать необходимую силу, мощность, скорость, выносливость и т. д. то есть, выполнить необходимую работу с заданными параметрами). На основе механизма рекрутинга строится механизм межмышечной и внутримышечной координации. Другими словами, для сложных двигательных актов ЦНС должна координировать не просто схему включения/выключения отдельных мышц и мышечных групп, но и последовательность включения/выключения отдельных групп волокон внутри самой мышцы. Т.е., точно координировать и регулировать работу всех мышечных пулов в каждой отдельно взятой мышце. И главное в данном процессе является выключение мышечных волокон и возможность мышцы расслабиться и быть растянутой до первоначальной или заданной программой длинны. Из физиологии мышечного сокращения известно, что утомление не только снижает сократительную способность мышцы (соответственно вызывая потерю в силе), но и способность вовремя расслабляться, что увеличивает погрешности при управлении движением.
Чем выше процентное соотношение медленно сокращающихся тонических волокон, тем выше способность мышцы к накоплению и отдаче эластической энергии [рекуперация]. Соответственно тем выше экономичность самого движения.
КПД мышц достигает 35%, что является хорошим показателем производительности по сравнению с другими типами «двигателей», но даже при таком условии расход метаболической энергии на выполнение мышечного сокращения очень высок. И требуется значительное количество времени на пополнение энергетического запаса и накопления резервов.
Любое движение может быть представлено как перемещение из одной равновесной точки, в которой мышечная активность минимальна, в другую.
С точки зрения биомеханики поддержание равновесия и одновременное управление движениями являются достаточно сложными задачами, поскольку тело человека представляет собой многозвенную биомеханическую систему, в которой наиболее массивное звено (корпус) расположено высоко над площадью опоры, размеры которой ограничены площадью опорной базы.
Между звеньями существует сложное динамическое взаимодействие. Движение в любом отдельно взятом звене приводит к необходимости вырабатывать корректирующие силовые моменты (мышечное напряжение) во всех суставах кинематической цепи.
Для реализации любой двигательной задачи в рычажно-маятниковом механизме ЦНС нужно преодолеть проблему степеней свободы — в каждом определенном отрезке времени и траектории движения сегмента обеспечить его стабильность или мобильность относительно другого подвижного или неподвижного сегмента или опоры. Фактор длительности и силы напряжения мышц играют основную роль.
Ключевой функцией ЦНС является контроль избыточного движения, избыточного напряжения, стремление к минимизации степеней свободы или ограничение числа занятых независимых элементов движения.
Другими словами: ЦНС должна вовремя замыкать и размыкать суставы и контролировать скорость и углы сгибания-разгибания. При этом общий центр масс (ОЦМ) должен постоянно находиться под контролем ЦНС с целью препятствования падению.
Каждая мышца имеет свою индивидуальную структуру и специализацию. Но, несмотря на это, любое движение выполняется группами мышц посредством ансамблевого взаимодействия как внутри групп мышц со сходным действием, так и согласованно с группами антагонистов — мышц с противоположным вектором действия. Антагонизм довольно условен. При выполнении движения антагонисты выполняют контролирующую, тормозящую или стабилизирующую функцию по отношению к агонистам — мышцам, выполняющим основное движение.
Для выполнения различных задач ЦНС группирует мышцы в динамические мышечные цепи. Такие временные «союзы» направлены для решения конкретной задачи и не являются неизменными анатомическими образованиями. Такое групповое взаимодействие может требоваться для динамической стабилизации туловища (конечностей) или выполнения задач, требующих сложной координации. Одни и те же мышцы могут входить в состав различных мышечных цепей. Участие отдельных мышечных цепей различно даже при выполнении отдельных фаз движения. Чаще всего проблемы возникают при движениях со сложной структурой, когда одна и та же мышечная группа или отдельная мышца работает как агонист — выполняет основное движение, и стабилизирует сустав (одновременно являясь неотъемлемой частью системы стабилизации). Пример — поясничная мышца при сгибании позвоночника передними волокнами сгибает корпус, а задними контролирует углы сгибания позвонков относительно друг друга — стабилизирует поясничный отдел позвоночника — препятствует сгибанию корпуса.
Контроль за фазами перехода концентрического сокращения в эксцентрическое и обратно требует повышенного внимания со стороны ЦНС и точного контроля над отдельными мышечными параметрами — скоростью, силой сокращения, временем расслабления. Такая работа ЦНС обозначается как внутримышечная и межмышечная координация. Для предотвращения перегрузки и истощения отдельных мышц и для оптимизации выполнения и повышения точности движения ЦНС использует синергии — содружественную работу мышц, повышающую их производительность с одновременным снижением рабочего износа и повышением общего коэффициента полезного действия мышечной группы.
При любом движении мышца может работать как агонист, антагонист, синергист. Смена фаз зависит от выполняемого движения. Причем фазы могут сменяться мгновенно.
ЦНС применяет синергии для обеспечения гибкости и вариабельности выполнения двигательной задачи. Также такой механизм позволяет предотвратить или снизить истощение метаболических резервов в каждой отдельно взятой мышце.
Наряду с управлением синергиями, ЦНС использует и другой механизм мышечной работы — коактивацию — одновременное напряжение агонистов-антагонистов — мышц с противоположным действием. Используется для динамической стабилизации суставов.
В норме одновременное напряжение мышц (коактивация) не превышает 2—3 (два-три) процента от максимального мышечного сокращения. Правомерно будет сказать, что благодаря конструкции скелета и свойствам мышечно-фасциальной системы передвижение осуществляется почти в холостом режиме благодаря упруго-эластическим свойствам мышц и возможности работать в режиме рекуперации энергии.
При маятниковом механизме, активация мышечного сокращения выполняет корригирующую функцию по возвращению ОЦМ в площадь опоры или используется при средних и интенсивных нагрузках непродолжительный период. Все остальное время мышечная система работает в фоновом тоническом режиме с кратковременными пиками повышения активности различных мышечных групп.
Причины мышечной боли
Разбор начну с боли, вызванной спазмом мышц, поскольку данный механизм заявлен как основной и этот «концепт» прочно вбит в головы терапевтам, семейным врачам, массажистам и мануальщикам. В последнее время в литературе для неврологов навязывается такой же примитивный взгляд на боли в спине, как и у терапевтов. Если и дальше так пойдет, то по уровню знаний отличить невролога от семейного врача будет невозможно. Привожу конкретный пример:
Практическая неврология: руководство для врачей под редакцией профессора А. С. Кадыкова (Серия «Библиотека врача-специалиста), 2011. Предназначено для врачей-неврологов, врачей общей практики, терапевтов, фельдшеров, студентов медицинских вузов, а также для всех специалистов, оказывающих помощь больным с заболеванием нервной системы.
Глава 11. Боли в спине. Авторы: Л. С. Манвелов, В. В. Шведков, А. С. Кадыков):
«Первоначально основной причиной возникновения болей в спине считали воспаление нервных корешков и стволов. В дальнейшем инфекционно-аллергическая теория патогенеза болей в спине постепенно стала уступать место вертеброгенной, чему в определенной степени способствовал успех операций по поводу грыжи диска. Одно время все боли в спине стали объяснять дегенеративно-дистрофическими изменениями позвоночника, сдавлением грыжей межпозвонкового диска нервного корешка. В этот период появляется и соответствующая терминология: дискогенный корешковый компрессионный синдром, вертеброгенная радикулопатия, вертеброгенный рефлекторный синдром.
В 1980–1990–е гг. у неврологов стала превалировать теория преимущественно мышечного происхождения болей в спине. В настоящее время считается, что почти 90% случаев причиной болей в спине являются миофасциальные синдромы, [выделено мной] а на долю вертеброгенных нарушений приходится не более 10%. Это отражает и соответствующая терминология (дорсалгия, люмбалгия, миофасциальный синдром).
Этиология и патогенез болей в спине.
По современным представлениям, чаще всего, причинами возникновения болей в спине являются:
— патологические изменения в позвоночнике, прежде всего, дегенеративно-дистрофические;
— патологические изменения в мышцах, чаще всего миофасциальный синдром;
— патологические изменения в органах брюшной полости;
— заболевания нервной системы.
Факторами риска возникновения болей в спине считаются:
— тяжелые физические нагрузки;
— неудобная рабочая поза;
— травма, охлаждение, сквозняки;
— злоупотребление алкоголем;
— депрессия и стрессы;
— профессиональные заболевания, связанные с воздействием высоких температур (в частности, в горячих цехах), лучевой энергии, резкими колебаниями температур, вибрации.
Среди вертебральных причин болей в спине различают:
— ишемию корешка (дискогенный корешковый синдром, дискогенная радикулопатия), которая возникает вследствие сдавления корешка грыжей диска;
— рефлекторные мышечные синдромы, причиной которых могут быть различные дегенеративно-дистрофические изменения в позвоночнике.
Определенную роль в возникновении болей в спине могут играть различные функциональные расстройства поясничного отдела позвоночника, когда из-за неправильной позы возникают блоки межпозвонковых суставов и нарушается их подвижность. В суставах, расположенных выше и ниже блока, развивается компенсаторная гипермобильность, приводящая к мышечному спазму».
Собственно «Миофасциальный синдром»:
Миофасциальный синдром является основной причиной развития болей в спине. Он может возникать вследствие перенапряжения (во время тяжелой физической нагрузки), перерастяжения и ушибов мышц, нефизиологичной позы во время работы, реакции на эмоциональный стресс, укорочения одной ноги и даже плоскостопия. Кроме того, к предрасполагающим факторам относятся:
— гиповитаминоз B1, B6, B12, фолиевой и аскорбиновой кислот;
— недостаточность микроэлементов (калия, кальция, магния, железа;
— гиподинамия;
— подагрический диатез;
— хронические инфекции;
— нарушение сна.
Аналогами термина «миофасциальный синдром» являются: миалгия миофиброзит, миофасцит.
Для миофасциального синдрома характерно наличие так называемых «курковых» зон (триггерных точек), надавливание на которые вызывает боль, часто иррадиирующие в соседние области.
Помимо миофасциального болевого синдрома причиной болей могут быть и воспалительные заболевания мышц — миозиты».
(Практическая неврология: руководство для врачей/под ред. проф. А. С. Кадыкова, Л. С. Манвелова, В. В. Шведкова. — М.: ГЭОТАР — Медиа, 2011. — 448 с. (Серия «Библиотека врача-специалиста)
Как мы видим, здесь все те же невнятные рассуждения о «дегенеративно-дистрофических изменениях», «миофасциальном синдроме», «спазме». Однозначно, по факту можно констатировать деградацию представлений о патогенезе остеохондроза позвоночника и выраженный откат в понимании такой сложной и многогранной патологии как боль в спине. На проблему примитивного представления будущих врачей о данной патологии можно было бы закрыть глаза [существуют другие медицинские пособия], если бы авторы данного пособия не совали свой нос в вопросы физической реабилитации. Глава 11 «Боли в спине» снабжена 4–мя комплексами упражнений при болях в спине. Я много видел разных комплексов, но таких позорных как в данном пособии — никогда. Первые три комплекса — это разорванный на три части комплекс из 9 упражнений проф. Шостака Н. А. и Правдюка Н. Г., (кафедра факультетской терапии РГМУ), но с «оторванным» упражнением №9 [подъем ног поочередно из положения лежа на животе]. Это единственное упражнение в комплексе Шостака на укрепление мышц спины. Мало того, что эти «уникумы» взяли чужой комплекс, так они умудрились найти самый бестолковый из всех возможных. (Комплекс Шостака–Правдюка будет рассматриваться в главе, посвященной применению упражнений). Вопрос, собственно, в другом: Манвелов, Шведков и Кадыков всю патологию позвоночника свели к «мышечным спазмам», «триггерным точкам» и «блокированию суставов позвоночника», поэтому весь предложенный ими [позаимствованный] комплекс состоит из упражнений на сгибание, скруток позвоночника, висов — т.е. упражнений для устранения «укороченных/спазмированных» мышц. Данные авторы не понимают, что остеохондроз позвоночника и кумулятивная травма связочной ткани суммарно составляют 3/4 всей патологии у пациентов, страдающих болью в спине, и данные две патологии сопровождаются нестабильностью (!) в ПДС. Какие скрутки, какие висы на межкомнатных дверях? Да сколько можно тиражировать эту глупость?
Врачи рекомендуют эти упражнения пациентам, которые самозабвенно пытаются растянуть «спазмированные мышцы», снять «блоки позвонков» и добиться «гибкости позвоночника» как рекомендуют авторы «для восстановления подвижности мышц», тем самым — добивая позвоночник и усугубляя течение заболевания.
«Спазм»
Начну со всеми любимой концепции «спазма».
Звучит она так: «боль вызывает спазм в мышце/мышцах, длительное напряжение вызывает ишемию в спазмированных мышцах, что приводит к накоплению продуктов обмена, которые раздражают хемочувствительные ноцицепторы, которые, в свою очередь, усиливая боль, приводят к еще более выраженному спазму». Данный «порочный круг» назван «Цикл „боль–спазм–боль“».
Данная гипотеза свое время была выдвинута Janet Travell et al (1942), (статья «Боль и нарушение функции в плече и руке»). С тех пор ее пытаются «задвинуть обратно», но безрезультатно.
Критический обзор работ (рассматривался период с 1941 по 1985 г.г.), посвященный данному вопросу, был сделан Roland M. O., (1986). Его выводы:
«Боль и спазм — частые клинические признаки пациентов с проблемами спины, и известно, что при определенных обстоятельствах они могут возникать независимо друг от друга. Например, у некоторых пациентов наблюдается выраженный мышечный спазм, но нет боли (например, некоторые пациенты с острой кривошеей). Так же встречаются пациенты, которые имеют сильную боль, но без спазма (например, при синдроме «гипермобильности»).
Между этими крайностями встречаются пациенты, у которых клинически выявлены как боль, так и спазм, и это может быть продемонстрировано в лабораторных условиях.
Ни одно из приведенных выше свидетельств не доказывает существования цикла боль-спазм-боль. В действительности, ряд отдельных исследований серьезно испорчены методологическими ошибками. Тем не менее, баланс доказательств указывает на то, что боль и спазм при заболеваниях позвоночника не всегда возникают самостоятельно [независимо]».
(Roland, M. O. (1986). A critical review of the evidence for a pain-spasm-pain cycle in spinal disorders. Clinical Biomechanics, 1 (2), 102–109).
Lund et al. (1991):
«Обзор изменений двигательных реакций, происходящих при хронических мышечных болях, убедил нас в том, что модель „порочного круга“ и другие аналогичные модели, основанные на принципе взаимного усиления боли и мышечной гиперактивности, — неверны».
(Lund, J. P., Donga, R., Widmer, C. G., & Stohler, C. S. (1991). The pain-adaptation model: a discussion of the relationship between chronic musculoskeletal pain and motor activity. Canadian Journal of Physiology and Pharmacology, 69 (5), 683–694. doi:10.1139/y91—102).
Сама Janet Travell, осознав свою ошибку, также попыталась опровергнуть свою же гипотезу, но, увы, ее попытка тоже не увенчалась успехом. Данная гипотеза настолько нравится фармакологам, что, несмотря на всеобщие усилия ученых, она [гипотеза] регулярно «оживляется» в различных книгах и монографиях, что основательно закрепило в сознании врачей ее как «единственно верную».
Мнение самой J.Travell по данному вопросу:
«Цикл «боль–спазм–боль». Устаревшая концепция цикла «боль–спазм–боль» не заслуживает экспериментальной проверки ни с точки зрения физиологии, ни с клинической точки зрения.
Физиологические исследования показали, что боль, исходящая из мышц, обладает тенденцией угнетать, но не облегчать рефлекторную сократительную активность этой мышцы. Walsh подробно объяснил, как эта ошибочная концепция подкреплялась неправильными представлениями о возникновении нормальных двигательных рефлексов у человека, основываясь на экспериментальных исследованиях спинного мозга у кошек, и каким образом такой ошибочной точке зрения удалось просуществовать в течение всего XX столетия.
В 1989г. Ernest Johson, редактор журнала «American Journal of Physical Medicine», привел ошеломляющее свидетельство того, что тесная взаимосвязь общего восприятия мышечной боли и мышечного спазма является мифом и что этот миф упорно поддерживался из коммерческих интересов.
Современная разновидность концепции «боль–спазм–боль» — «теория активной стрессовой боли» также несостоятельна по тем же причинам».
(Трэвел и Симонс. Миофасциальные боли и дисфункции. Руководство по триггерным точкам в 2 томах. Издание 2-е, переработанное и дополненное. Москва «Медицина» 2005).
Все доходчиво и без лишних сентиментов.
Концепцию «боль–спазм–боль» версии J.Travell мы разбирать не будем в силу вышезаявленного.
Для нас интересна другая распространенная «версия» данной концепции, а именно версия Johansson and Sojka (1991).
(Johansson, H., & Sojka, P. (1991). Pathophysiological mechanisms involved in genesis and spread of muscular tension in occupational muscle pain and in chronic musculoskeletal pain syndromes: a hypothesis. Medical hypotheses, 35 (3), 196—203).
По непонятным мне причинам данную гипотезу также отнесли к концепции «Боль-спазм–боль», хотя в оригинальной статье о спазме было упомянуто только в конце статьи и только при упоминании гипотезы J.Travell. Вероятно, это связано с тем, что авторы также употребляли выражение «порочный круг» как и J.Travell.
В оригинальной гипотезе Johansson and Sojka описывался предполагаемый механизм, лежащий в основе «Синдрома хронической мышечно-скелетной боли» и речь шла не о мышечном спазме, а о повышении мышечного тонуса, вызванного повышением активности в гамма- петле. И авторы употребляли выражение типа «ригидность (жесткость) мышц» (muscle stiffness), а не «мышечный спазм» (muscle spasm).
Разберем данную гипотезу, чтобы понять где, в очередной раз, фармакологическое лобби нас ловко одурачило.
Активные «участники» процесса под названием «спазм» (правильно говорить «мышечная ригидность»:
— Ноцицепторы А-дельта (III тип) и С (IV тип);
— Моторная единица — альфа-мотонейрон и иннервируемые им экстрафузальные волокна;
— Гамма–мотонейроны (динамический и статический) и иннервируемые ими интрафузальные волокна, управляющие чувствительностью «веретена» — датчиками скорости растяжения мышцы и ее текущей длинны;
— Механорецепторы кожи, суставов, связочного аппарата.
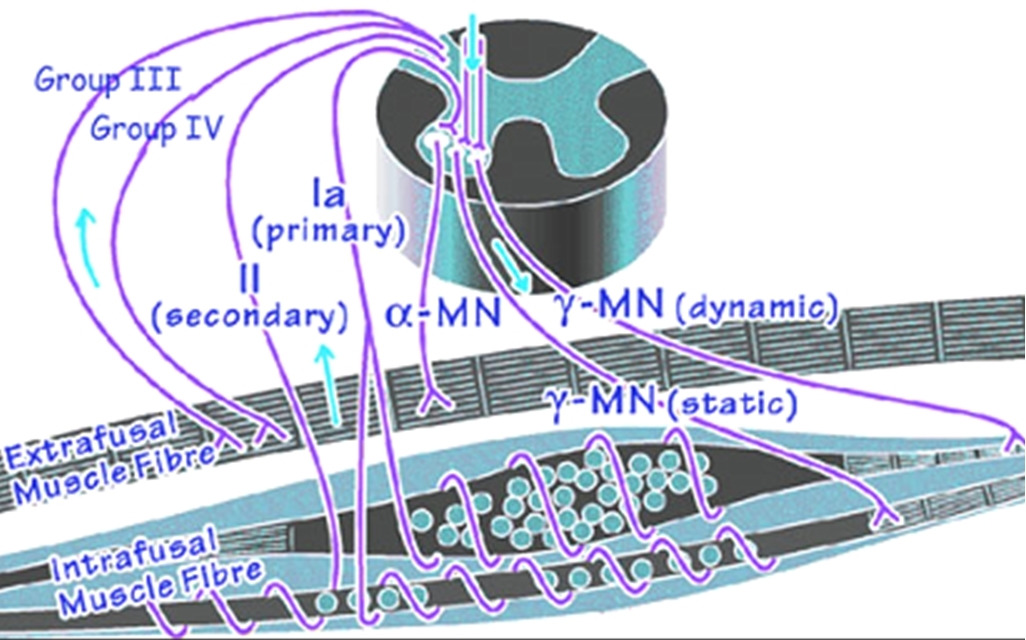
Некоторые пояснения. Мышечные волокна в постуральных мышцах никогда не бывают полностью расслаблены, в них всегда сохраняется некоторое напряжение. Вызывается единичными разрядами альфа-мотонейронов.
Выделяют упругоэластический и сократительный тонус мышц. В расслабленной мышце фиксируется только упругоэластический тонус, определяемый физическими свойствами соединительной ткани. Электромиографически мышца «молчит».
Мышечный тонус — длительное напряжение скелетных мышц, отличающееся малой утомляемостью и большой экономичностью. За регулировку тонуса мышц на сегментарном уровне отвечает гамма-петля, являющаяся частью сервомеханизма альфа-гамма сопряжения.
Управление движением через гамма-петлю является одним из экономичных и автоматизированных процессов мышечного сокращения. Когда человек обучается какому-либо новому «мышечному» навыку, тогда структуры мозга, отвечающие за движение, контролируют мышечное сокращение непосредственно активируя альфа-мотонейроны, т.е. постоянно «присматривают» за их возбуждением и сокращением мышц. Прямой путь контроля над выполнением двигательных программ очень трудоемкий и энергозатратный для мозга процесс и используется только в начале обучения движению. По мере усвоения нового моторного навыка, ЦНС передает обязанности контроля над выполнением движения в подкорковые структуры и двигательные центры спинного мозга. По мере автоматизации движения управление происходит уже через гамма–петлю — заблаговременную преднастройку параметров мышцы на основе накопленных в «моторной» памяти шаблонов, заготовок и целых программ.
Принцип работы гамма-петли и поддержания мышечного тонуса:
от гамма-мотонейронов поступает активизирующая импульсация к интрафузальным мышечным волокнам, вызывая их сокращение, что приводит к растяжению экваториальной части рецептора, снабженного чувствительными волокнами. Это вызывает возникновение ритмичной биоэлектрической активности, которая по афферентной (чувствительной) части рефлекторной дуги проводится к малым (тоническим) альфа-мотонейронам спинного мозга и их активации. В них возникают продолжительные синхронные разряды (импульсы), которые по аксонам этих клеток направляются к экстрафузальным мышечным волокнам, вызывая их продолжительное сокращение, необходимое для поддержания мышечного тонуса.
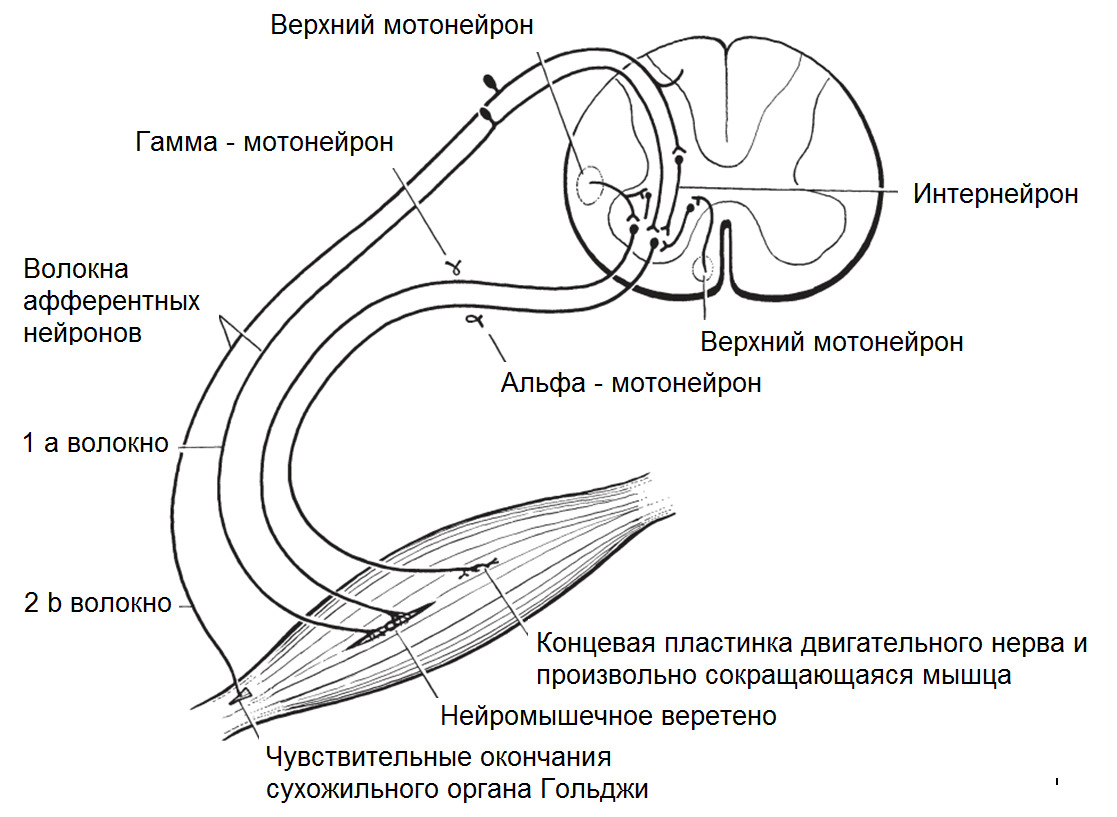
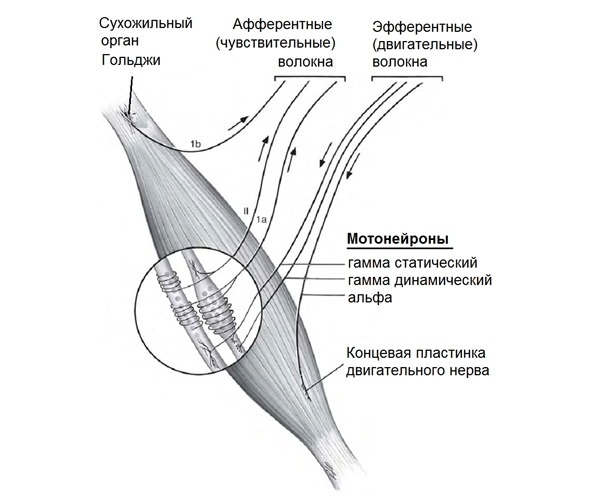
Гамма-петля состоит из двух компонентов — тонического (II) и фазического (динамический) (Ia).
Тонический отвечает за передачу данных на моторные (двигательные) нейроны о текущей длине мышцы.
Фазический — о скорости растяжения/сокращения мышцы.
Мышечный тонус в управлении движением:
— обеспечивает исходное положение тела (звена), необходимое для осуществления любого целевого движения в гравитационном поле земли;
— определяет физиологические параметры мышц–эффекторов: начальную длину, механические (упруго-вязкие) свойства;
— облегчает управление, устраняя (или ограничивая) лишние степени свободы.
Вызвать сокращение мышцы можно двумя путями: подав прямой сигнал на альфа-мотонейроны с последующим сокращением подконтрольных мышечных волокон или подав сигнал на гамма-мотонейроны, активировав рефлекс растяжения, что вызовет сокращение мышечных волокон (или мышцы в целом). При сложнокоординированных движениях сигнал подается на три нейрона сразу: альфа, бета и гамма.
В теории гипотезы Johansson and Sojka заявлено: «Синдром хронической мышечно-скелетной боли» развивается в мышцах, регулярно задействованных в выполнении статической работы или работающих в режиме монотонных сокращений. Данный тип боли устойчив к лечению и склонен к переходу в хроническую.
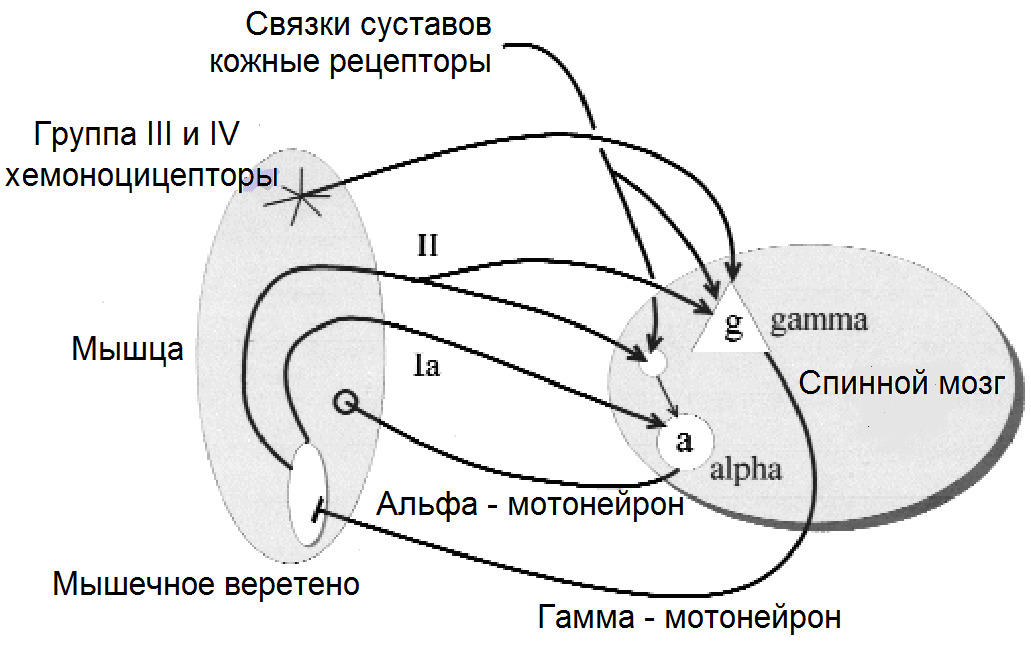
Длительная работа в статическом положении или длительное сокращение мышцы приводит к накоплению метаболитов (не выведенных в кровоток продуктов обмена). Сопутствующее микроповреждение связочной и мышечной ткани также приводит к выработке медиаторов воспаления. Данный «коктейль» активирует ноцицепторы А-дельта и С волокон, реагирующие на химический стимул. Волокна ноцицептивных нейронов контактируют с гамма-мотонейронами спинного мозга, возбуждая их, что приводит к сокращению интрафузальных мышечных волокон и активации чувствительных волокон внутри «веретена», что, в свою очередь, приводит к «разрядам» в первичном (динамический, тип Ia) и вторичном (статический, тип II) волокнах. Чувствительные нейроны группы Ia напрямую контактируют с альфа-мотонейронами; чувствительные нейроны группы II контактируют с альфа-мотонейронами через интернейрон (посредник) и напрямую с гамма-мотонейронами. Соответственно, возбуждающий сигнал, приходящий по чувствительным волокнам (Ia и II), возбуждает как альфа, — так и гамма-мотонейроны, что приводит к сокращению иннервируемых ими волокон и повышению тонуса мышц. Длительная статическая работа (или монотонная) на фоне общего повышения тонуса в мышцах приводит к более быстрому накоплению продуктов обмена и более ранней активации ноцицепторов (усугубляет ситуацию через ишемию ткани). Длительная стимуляция гамма-мотонейронов (через волокна чувствительного гамма нейрона тип II) создает второй добавочный «порочный круг» за счет повышения чувствительности рефлекса растяжения. Данный тип уже не зависит от активации ноцицепторов и повышенный тонус воспроизводится рефлекторно.
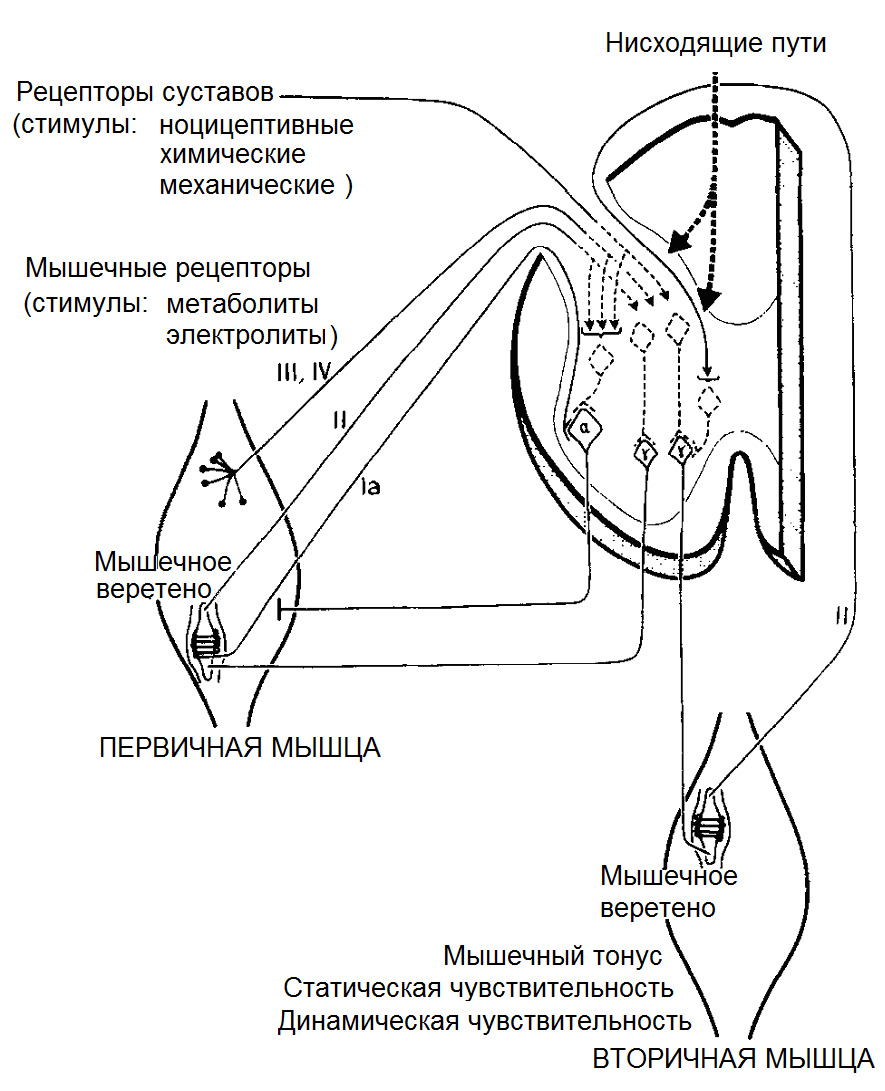
Достаточно легкого неконтролируемого движения или рывка, и рефлекс растяжения мгновенно активирует мышечное сокращение. Чем сильнее «бомбежка» гамма-мотонейронов ноцицептивными («болевыми») или механорецепторами связочного аппарата, тем больше шансов на «расползание» возбуждения по всему сегменту и вовлечение в патологический процесс соседних мышц. Малые мотонейроны, иннервирующие тонические волокна, имеют очень низкий порог активации. Соответственно, чтобы их активировать достаточно и слабых, но продолжительных стимулов. Сильное бомбардирование мото-нейронов разного типа может вызвать выраженное локальное мышечное напряжение, если сила и частота «бомбардировки» превысит величину порога активации более крупных мотонейронов, иннервирующих более крупные моторные единицы».
Что не так в данной гипотезе и почему ее необоснованно отнесли в категорию «боль-спазм–боль»?
Чтобы ответить на этот вопрос для большей наглядности и информативности воспользуюсь принципиальной схемой элементарной моторной «функциональной единицы» сегмента спинного мозга, относящейся по классификации Н. А. Бернштейна к уровню организации движения «А». Данная схема хорошо известна специалистам по реабилитации, поскольку ее предложил Siegfried Mense (Сигфрид Менс), хорошо известный своими книгами «Мышечная боль: понимание механизмов» (2010) (Muscle Pain: Understanding the Mechanisms) и «Мышечная боль: диагностика и лечение» (2010) (Muscle Pain: Diagnosis and Treatment).
Напомню за что отвечает данный уровень:
А — Уровень тонуса (руброспинальный) — организация движений в поле земного тяготения.
Уровень определяет функциональные возможности мускулатуры через преднастройку мышечного тонуса, влияя на сегментарный аппарат спинного мозга и фоновое изменение его возбудимости. Отвечает за реципрокную (взаимозависимую, взаимовлияющую) иннервацию мышц, регулирует силовые и скоростные характеристики сокращения мышц. Абсолютный монополист по тонусу мышц во всей центральной нервной системе.
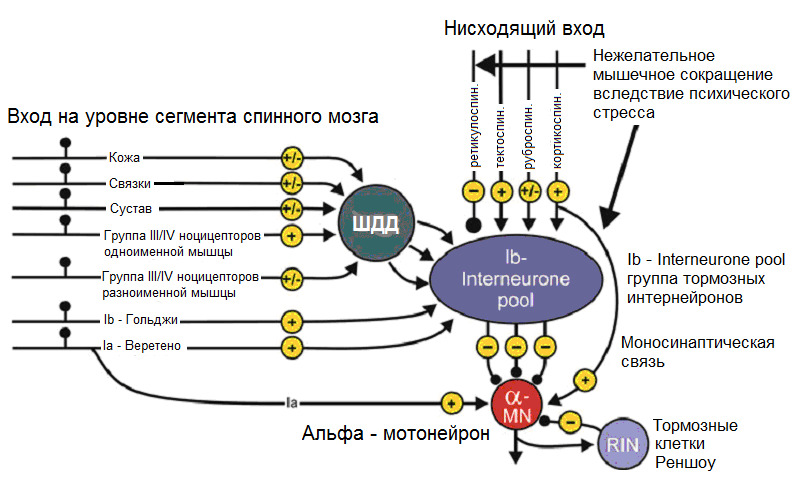
На данной схеме изображен «общий конечный путь» — альфа-мотонейрон, иннервирующий экстрафузальные мышечные волокна (собственно, то, что обыватель подразумевает под словом «мышца»). Данной схемой удобно пользоваться для объяснения всех феноменов, присущих мышечному сокращению и простой мышечной патологии. Знак «+» означает активацию альфа-мотонейрона или усиление действия группы интернейронов; соответственно, «-» означает торможение или ослабление действия. То есть, «плюс» на «минус» дает «усиленный минус» — усиление торможения альфа-мотонейрона, «минус» на «минус» дает «ослабленный минус» — ослабление торможения.
В учебниках по неврологии «общий конечный путь» чаще всего изображают в проекции на сегмент спинного мозга. Но это не принципиально. Смысл тот же.
Бесплатный фрагмент закончился.
Купите книгу, чтобы продолжить чтение.